[Most Recent Entries] [Calendar View]
Saturday, July 21st, 2018
| Time | Event | ||||||||||
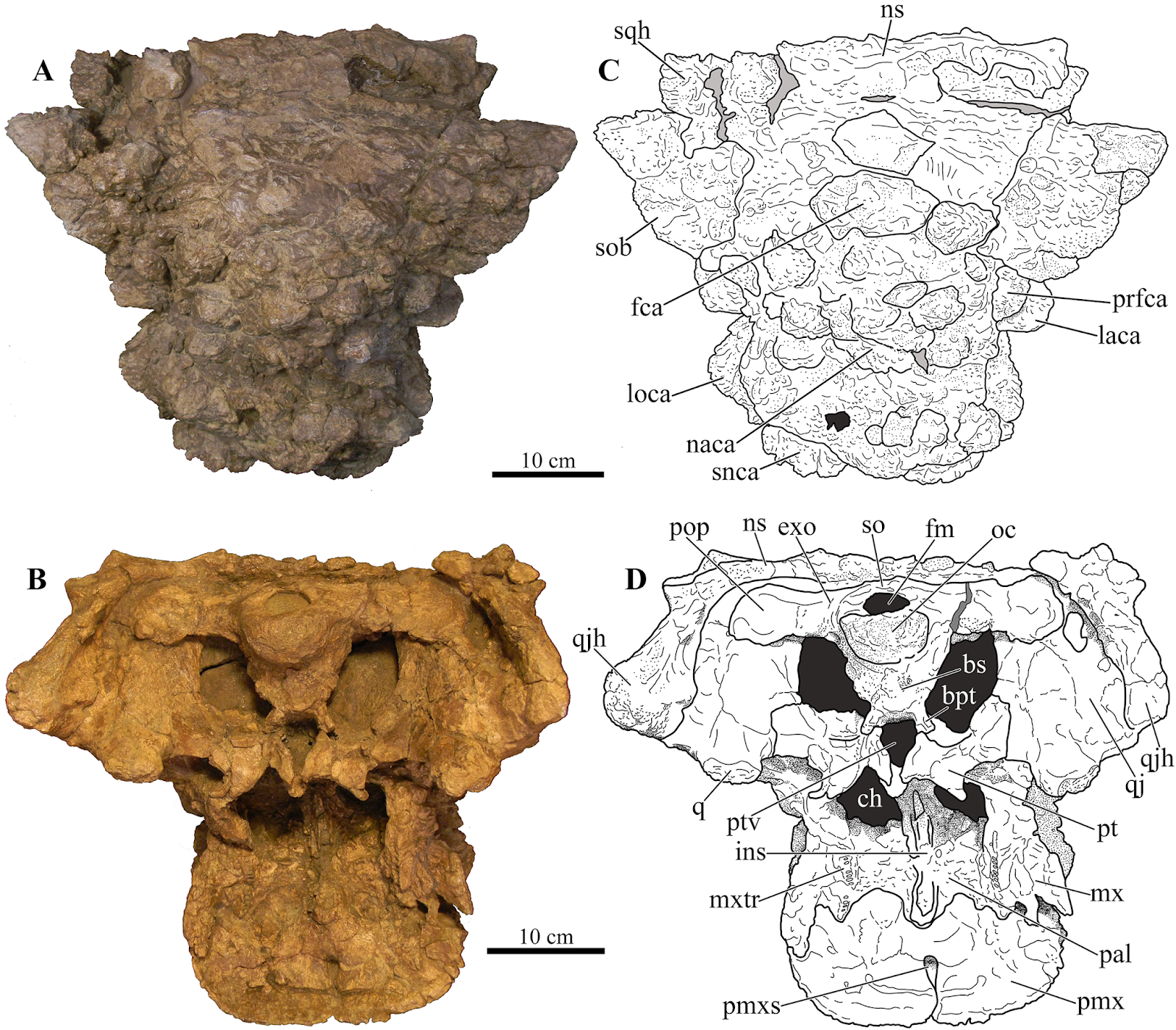
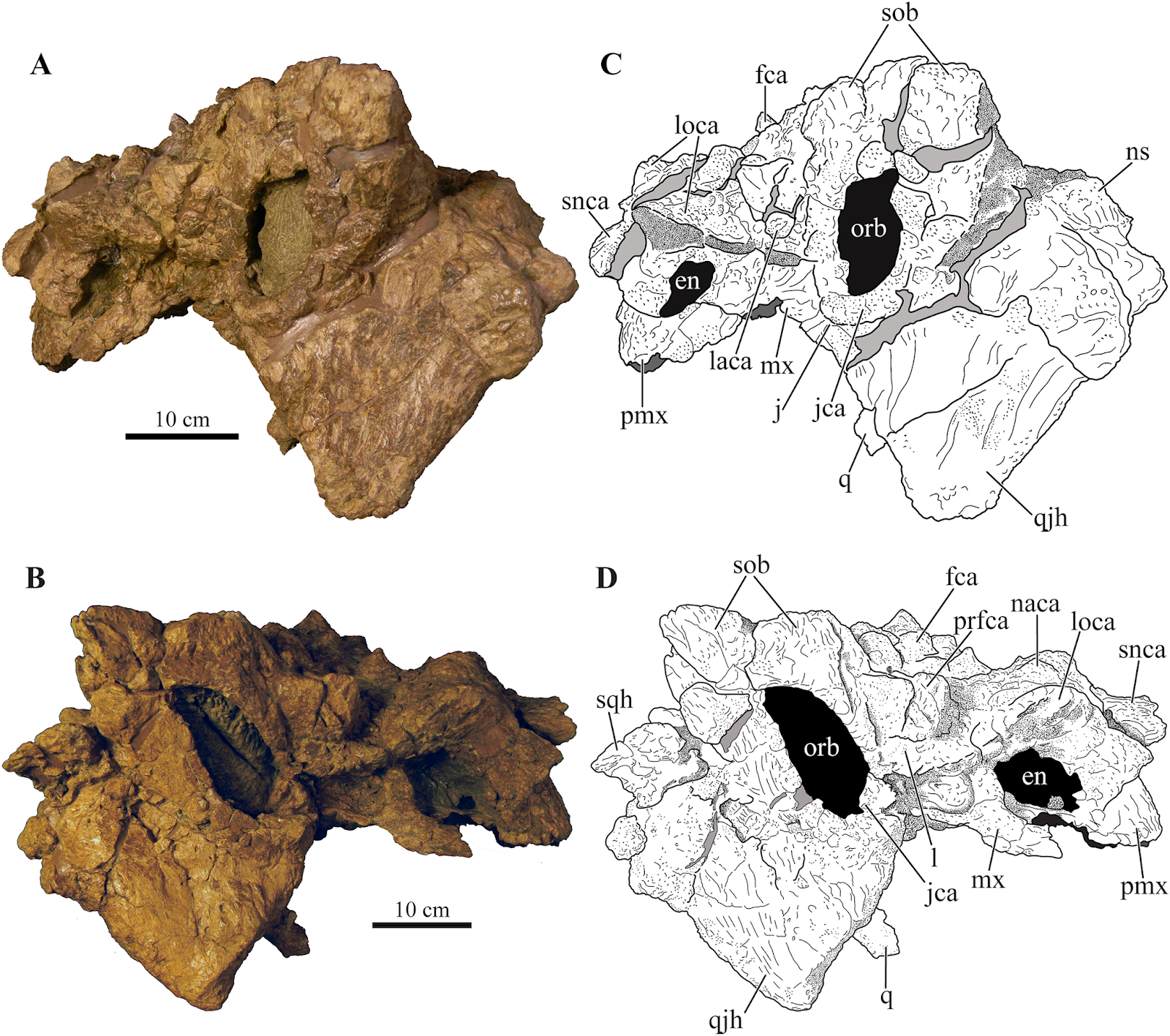
| 2:24p | [Paleontology • 2018] Akainacephalus johnsoni • A New southern Laramidian Ankylosaurid from the upper Campanian Kaiparowits Formation of southern Utah, USA Abstract A partial ankylosaurid skeleton from the upper Campanian Kaiparowits Formation of southern Utah is recognized as a new taxon, Akainacephalus johnsoni, gen. et sp. nov. The new taxon documents the first record of an associated ankylosaurid skull and postcranial skeleton from the Kaiparowits Formation. Preserved material includes a complete skull, much of the vertebral column, including a complete tail club, a nearly complete synsacrum, several fore- and hind limb elements, and a suite of postcranial osteoderms, making Akainacephalus johnsoni the most complete ankylosaurid from the Late Cretaceous of southern Laramidia. Arrangement and morphology of cranial ornamentation in Akainacephalus johnsoni is strikingly similar to Nodocephalosaurus kirtlandensis and some Asian ankylosaurids (e.g., Saichania chulsanensis, Pinacosaurus grangeri, and Minotaurasaurus ramachandrani); the cranium is densely ornamented with symmetrically arranged and distinctly raised ossified caputegulae which are predominantly distributed across the dorsal and dorsolateral regions of the nasals, frontals, and orbitals. Cranial caputegulae display smooth surface textures with minor pitting and possess a distinct conical to pyramidal morphology which terminates in a sharp apex. Character analysis suggests a close phylogenetic relationship with N. kirtlandensis, M. ramachandrani, Tarchia teresae, and S. chulsanensis, rather than with Late Cretaceous northern Laramidian ankylosaurids (e.g., Euoplocephalus tutus, Anodontosaurus lambei, and Ankylosaurus magniventris). These new data are consistent with evidence for distinct northern and southern biogeographic provinces in Laramidia during the late Campanian. The addition of this new ankylosaurid taxon from southern Utah enhances our understanding of ankylosaurid diversity and evolutionary relationships. Potential implications for the geographical distribution of Late Cretaceous ankylosaurid dinosaurs throughout the Western Interior suggest multiple time-transgressive biogeographic dispersal events from Asia into Laramidia.
 Systematic paleontology Ornithischia Seeley, 1887 sensu Padian and May, 1993 Thyreophora Nopcsa, 1915 sensu Sereno, 1986 Ankylosauria Osborn, 1923 sensu Carpenter, 1997 Ankylosauridae Brown, 1908 sensu Sereno, 1998 Ankylosaurinae Brown, 1908 sensu Sereno, 1986 Ankylosaurini Arbour and Currie, 2016 Akainacephalus, gen. nov. Akainacephalus johnsoni, sp. nov.
Holotype: UMNH VP 20202, a partial skeleton comprising a complete skull, both mandibles, predentary, four dorsal, four dorsosacral, three sacral, one caudosacral, and eight caudal vertebrae, dorsal ribs, a complete tail club, both scapulae, left coracoid, right humerus, right ulna, partial left ilium, left femur, left tibia, left fibula, phalanx, two partial cervical osteoderm half rings, and 17 dorsal and lateral osteoderms of various sizes and morphologies. Type locality: UMNH VP Locality 1109 (“HMG Quarry”), Horse Mountain area, GSENM, Kane County, southern Utah, USA. Type stratigraphic horizon and age: UMNH VP Locality 1109 is a multitaxic bonebed deposited in a crevasse splay sandstone within the lower portion of the middle unit of the upper Campanian Kaiparowits Formation (Fig. 2A). The stratigraphic position of this site is approximately 190 m from the base of the formation (Roberts et al., 2013: fig. 6.3) and within approximately one meter stratigraphic proximity of the recently dated bentonite ash bed KP-07, which has produced a U-Pb zircon age of 76.26 ± 0.10 Ma (Roberts et al., 2013), providing a precise age constraint for Akainacephalus johnsoni. Etymology: The genus name is derived from the Greek akaina, meaning “thorn” or “spine,” referring to the thorn-like cranial caputegulae of the holotype; and “cephalus,” the Greek meaning for head. The specific epithet honors Randy Johnson, volunteer preparator at the Natural History Museum of Utah, who skillfully prepared the skull and lower jaws of UMNH VP 20202. Diagnosis: Akainacephalus johnsoni possesses the following autapomorphies: massive supraorbital bosses in lateral view, forming a tall backswept flange extending laterally over the orbits, and enveloping the anterodorsal and posterior margins of the orbit; nearly vertical projecting triangular quadratojugal horns; frontal possesses a large, flat, and centrally positioned hexagonal-shaped caputegulum; a combination of tightly spaced, symmetrically positioned pyramidal and conical-shaped caputegulae across the frontonasal region; a distinct midline row of conical-shaped caputegulae across the nasal region, symmetrically separating caputegulae situated dorsolaterally; basioccipital foramen anterior and dorsally to the occipital condyle. A. johnsoni also possesses a unique combination of character states: shares with Nodocephalosaurus kirtlandensis the presence of a large, laterally oriented supranarial osteoderm forming the postmaxillary/lacrimal ridge dorsal to the external nares; differs from Tsagantegia longicranialis, Talarurus plicatospineus, Pinacosaurus grangeri, all northern Laramidian taxa and Ziapelta sanjuanensis but shares with Nodocephalosaurus kirtlandensis, Minotaurasaurus ramachandrani, Saichania chulsanensis, and Tarchia kielanae the presence of well-pronounced cranial ornamentation located along the nasal and frontal regions of the skull that are characterized by a dense array of well-defined caputegulae with a distinct conical (N. kirtlandensis) and pyramidal (M. ramachandrani, S. chulsanensis, T. kilanae) morphology; shares with Euoplocephalus and Zuul crurivastator a globular surface texture on the tail club knob, which differs from the smoother texture in Ankylosaurus magniventris; differs from ZPAL MgD I/113, cf. Pinacosaurus, Saichania chulsanensis, and Dyoplosaurus acutosquameus, but similar to Anodontosaurus lambei, Euoplocephalus tutus, Zuul crurivastator, and Ankylosaurus magniventris in having a wider than long tail club knob ratio; and shares with ZPAL MgD I/113, cf. Pinacosaurus, D. acutosquameus, and Zuul crurivastator triangular osteoderms along the lateral surfaces on the proximal portion of the tail.

  Conclusion: Akainacephalus johnsoni (UMNH VP 20202) is a new taxon of ankylosaurid dinosaur from the upper Campanian Kaiparowits Formation of southern Utah, USA. It consists of a complete cranium and significant amount of diagnostic postcranial material that can be distinguished from all other known Late Cretaceous Laramidian ankylosaurids. Akainacephalus johnsoni is closely related to its stratigraphically younger sister taxon, Nodocephalosaurus kirtlandensis from the Kirtland Formation of New Mexico. Both taxa are more closely related to Asian ankylosaurids than they are to other Laramidian ankylosaurids. This suggests multiple ankylosaurid dispersal events from Asia to Laramidia during the Late Cretaceous. Lastly, together with Dyoplosaurus acutossquameus and Scolosaurus cutleri (∼77 Ma) from northern Laramidia, Akainacephalus johnsoni represents one of the older known ankylosaurid dinosaurs (∼76.3 Ma) from the Late Cretaceous of western North America. Jelle P. Wiersma and Randall B. Irmis. 2018. A New southern Laramidian Ankylosaurid, Akainacephalus johnsoni gen. et sp. nov., from the upper Campanian Kaiparowits Formation of southern Utah, USA. PeerJ. 6:e5016. DOI: 10.7717/peerj.5016 Newly discovered armored dinosaur from Utah reveals intriguing family history phys.org/news/2018-07-newly-armored-dino Mystery of Spiky-Skulled Utah Dinosaur Solved livescience.com/63108-newfound-ankylosau    | ||||||||||
| 2:42p | [Herpetology • 2018] Boana icamiaba • Phylogeny of Map Tree Frogs, Boana semilineata Species Group (Anura: Hylidae), with A New Amazonian Species
Abstract. Gladiator Frogs (Boana) is a Neotropical group comprised of 92 species sorted into seven species groups. Herein, we present a phylogeny of the Boana semilineata species group, including all valid nominal species currently or suspected to be assigned to it— many sequenced for the first time. Parsimony and maximum likelihood analyses of two genes (16S and COI) did not support the monophyly of the B. semilineata group as currently defined. To remedy this, B. secedens, currently assigned to the B. puchella species group, is transferred to the B. semilineata group. On the other hand, our analyses corroborate the inclusion of B. hutchinsi, B. pombali and B. wavrini in the B. semilineata group. The reinterpretation of the morphology of the lower palpebral membrane in B. pombali (previously described as absent, but actually present) corroborates that the reticulated membrane is a synapomorphy of this species group (albeit homoplastic—present in a few species not assigned to this group). A recent paper suggested (based on the analyses of 16S alone) that at least six unnamed taxa (candidate species) are hidden behind the names B. geographica and B. semilineata. Our analyses resulted in the discovery of additional unnamed lineages, one of which is described and named herein. This new species is diagnosed, among other characters, by having the flanks, anterior and posterior surfaces of thigh with dark brown or blackish stripes, and by the presence of a prepollex forming a projecting spine. Keywords: Amazonia, Biodiversity, Cophomantinae, Hyla geographica, Taxonomy
Boana icamiaba sp. nov. Etymology: A noun in apposition, given as homage in memory of the legendary Amazonian female warriors, the Icamiabas. These warriors, first described by friar Gaspar de Carvajal (a member of the Francisco Orellana expedition; Medina, 1894), were members of isolated tribes composed of women only. So far as we are aware this is the second taxon to honor to the icamiabas, the other one being a hooded tickspider, Cryptocellus icamiabas Tourinho and Azevedo, 2007.  Pedro L.V. Peloso, Renan M. De Oliveira, Marcelo J. Sturaro, Miguel T. Rodrigues, Geraldo R. Lima-Filho, Youszef O.C. Bitar, Ward C. Wheeler and Alexandre Aleixo. 2018. Phylogeny of Map Tree Frogs, Boana semilineata Species Group, with a New Amazonian Species (Anura: Hylidae). South American Journal of Herpetology. 13(2); 150-169. DOI: 10.2994/SAJH-D-17-00037.1 New Frog Species Named After Fabled Female Warriors on.NatGeo.com/2muqL1k via @NatGeo | ||||||||||
| 3:06p | [Herpetology • 2018] Computational Molecular Species Delimitation and Taxonomic Revision of the Gecko Genus Ebenavia Boettger, 1878
Abstract Cryptic species have been detected in many groups of organisms and must be assumed to make up a significant portion of global biodiversity. We study geckos of the Ebenavia inunguis complex from Madagascar and surrounding islands and use species delimitation algorithms (GMYC, BOLD, BPP), COI barcode divergence, diagnostic codon indels in the nuclear marker PRLR, diagnostic categorical morphological characters, and significant differences in continuous morphological characters for its taxonomic revision. BPP yielded ≥ 10 operational taxonomic units, whereas GMYC (≥ 27) and BOLD (26) suggested substantial oversplitting. In consequnce, we resurrect Ebenavia boettgeri Boulenger 1885 and describe Ebenavia tuelinae sp. nov., Ebenavia safari sp. nov., and Ebenavia robusta sp. nov., increasing the number of recognised species in Ebenavia from two to six. Further lineages of Ebenavia retrieved by BPP may warrant species or subspecies status, but further taxonomic conclusions are postponed until more data become available. Finally, we present an identification key to the genus Ebenavia, provide an updated distribution map, and discuss the diagnostic values of computational species delimitation as well as morphological and molecular diagnostic characters. Keywords: BOLD, Operational Taxonomic Unit, Madagascar clawless gecko, Integrative taxonomy, Taxonomic inflation, Species complex Genus Ebenavia Boettger, 1878 Type species: Ebenavia inunguis Boettger, 1878 Diagnosis and description: A comprehensive diagnosis and description of the genus Ebenavia are provided in Nussbaum and Raxworthy (1998). Our data on the genus leads us to agree with the findings of that study, except that we were not able to confirm the presence of claws on the pes of any of the females we examined, as reported by these authors, in agreement with Boettger (1878) and many subsequent authors. We did not examine any specimens of E. maintimainty. Content: Ebenavia inunguis Boettger, 1878; E. boettgeri Boulenger, 1885; E. maintimainty Nussbaum & Raxworthy, 1998; and three new species described below. Distribution: Madagascar except most of the central and western areas; all major island areas of the Comoros Archipelago; Pemba Island (Tanzania); Mauritius. Ebenavia inunguis Boettger, 1878 Ebenavia boettgeri Boulenger, 1885, bona species Ebenavia tuelinae sp. nov. Etymology: A matronym dedicated to the first author’s partner Tülin (alternative spelling Tuelin) for her ceaseless support of this and other works and for her excellent spotting abilities in the field.
Ebenavia safari sp. nov. Etymology: ‘Safari’ means ‘voyage’ in the Kiswahili and Comoran (Shimaoré) languages spoken across the range of this species outside Madagascar. The name was chosen because this species dispersed over surprisingly long distances across the open ocean. It is treated as an unlatinised, invariable noun in apposition. Ebenavia robusta sp. nov. Etymology: The specific name is the feminine form of the Latin adjective ‘robustus’, meaning ‘robust’ or ‘sturdy’. It was chosen because this species is the largest and most sturdily built member of this genus of small geckos. Oliver Hawlitschek, Mark D. Scherz, Bernhard Ruthensteiner, Angelica Crottini and Frank Glaw. 2018. Computational Molecular Species Delimitation and Taxonomic Revision of the Gecko Genus Ebenavia Boettger, 1878. The Science of Nature. 105:49. DOI: 10.1007/s00114-018-1574-9 |









| << Previous Day |
2018/07/21 [Calendar] |
Next Day >> |