[Most Recent Entries] [Calendar View]
Thursday, March 21st, 2019
| Time | Event | ||||||||||||||
| 10:20a | [Arachnida • 2019] Surazomus saturninoae • A New Species of Surazomus (Schizomida) from eastern Amazon, with Comments on Homology of Male Flagellum and Mating March Anchorage in the Genus
Abstract Surazomus saturninoae sp. nov. is described from eastern Amazon. The male has a pentagonal flagellum, similar to those of three other species in the genus. These four species are herein gathered as the arboreus-group of Surazomus. We present a brief synopsis of chaetotaxy description in hubbardiines and several homology proposals for the flagellum of the species in the arboreus-group: the posterior lobes may be homologous to the lateral lobes of hubbardiine species with trilobed flagella; the setal brush with 4–5 setae on the posterior lobe may be composed of one Dl2 seta and enlarged lobular microsetae; the single, median posterior coupling pocket may be homologous to the pair of posterior pockets seen in other hubbardiines; the single, median anterior coupling pocket may be homologous to the pair of pockets on the anterior border of the flagellum seen in other hubbardiines. Based on the morphology of these pockets and the chelicerae within Surazomus, we discuss the anchoring mechanism during the mating march.
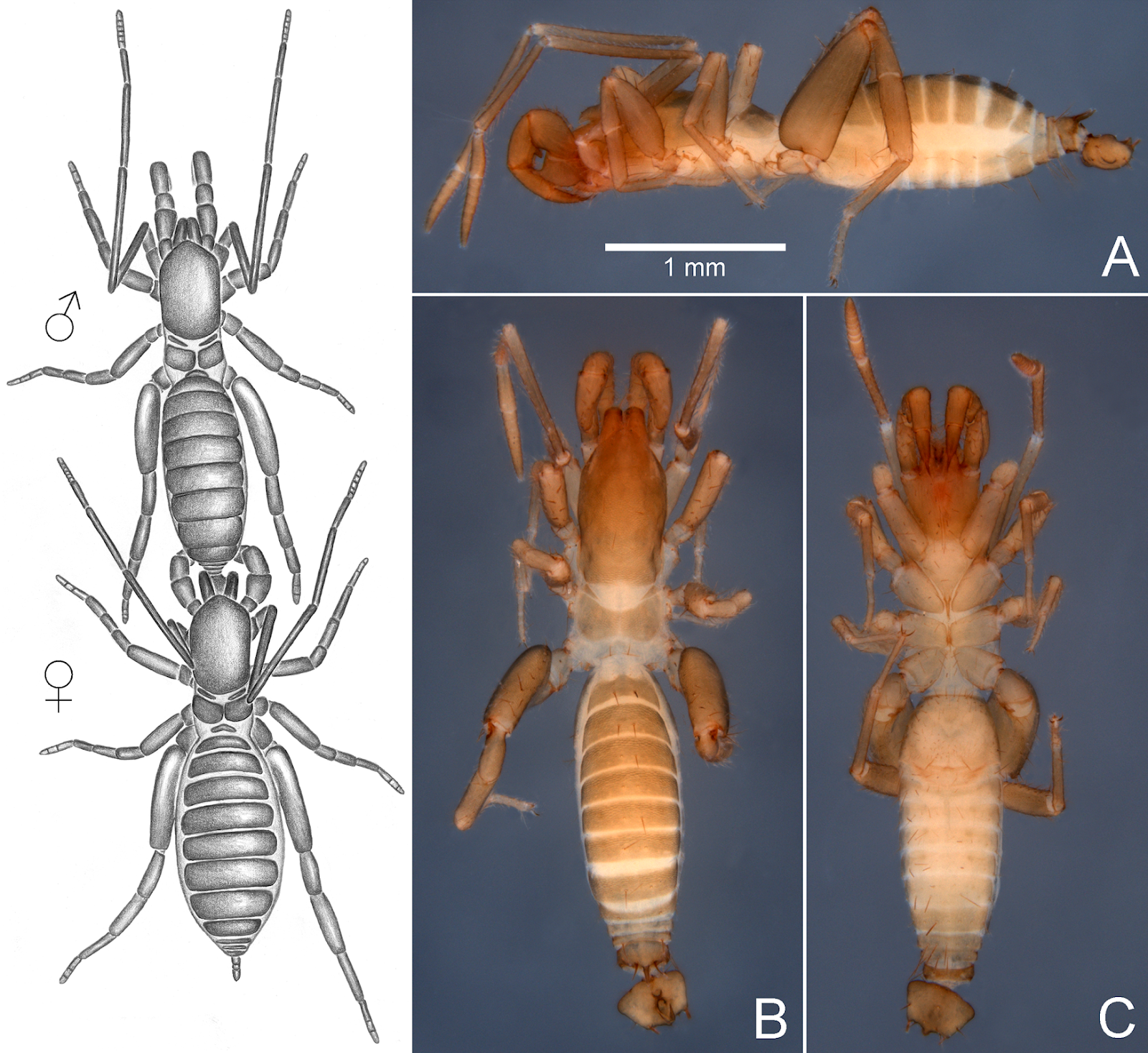
The arboreus-group of Surazomus. Within Surazomus, four species share features believed to be apomorphic and compose a species group, easily distinguished from the remaining species of the genus and traditionally recognized, but unnamed. To ease the discussion below, we herein propose the use of the informal name “the arboreus-group of Surazomus” to refer to that group. List of species of the arboreus-group: Surazomus arboreus Cokendolpher & Reddell, 2000; Surazomus manaus Cokendolpher & Reddell, 2000; Surazomus paitit Bonaldo & Pinto-da-Rocha, 2007; and Surazomus saturninoae sp. nov. Diagnosis. Species of the arboreus-group of Surazomus may be recognized by the males having pentagonal flagellum and bearing only two dorsal coupling pockets: one in front of Dm1 (AP), and another between Dm1 and Dm4 (PP), and by having a single large posterodorsal process on tergite XII (see Cokendolpher & Reddell [2000]: figs 11, 13, 20 and 23; Bonaldo & Pinto-da-Rocha [2007]: figs 2 and 3). Also, females have two pairs of slender, elongated spermathecal lobes with small bulbs followed by terminal constrictions (see Cokendolpher & Reddell [2000]: figs 14, 15, 24 and 25). Surazomus saturninoae sp. nov. Diagnosis from other species of the arboreus-group. Male: the overall shape of the flagellum of S. saturninoae sp. nov. is similar to that of S. arboreus, but the flagellum is as wide as long in S. arboreus (see Cokendolpher & Reddell [2000]: figs 11–13), while it is wider than long in S. saturninoae (Fig 6A). Also, the posterior dorsal hood (or dorso-median eminence) of S. arboreus is far from the posterior border of the flagellum, while it is close to the border in S. saturninoae (Fig 9D, in red). The widest portion of the flagellum in S. paitit is close to its middle length (see Bonaldo & Pinto-da-Rocha [24]: figs 2–4), while the widest portion of the flagellum in S. saturninoae is near its anterior border (Fig 6A). The posterodorsal process of abdominal segment XII of S. manaus (see Cokendolpher & Reddell [2000]: figs 20–23) and S. saturninoae (Fig 6A and 6C) is long, but the flagellum of S. manaus has parallel sides with the posterior lobes much longer than the flagellum body, while the flagellum in S. saturninoae has oblique sides and shorter posterior lobes. Female is unknown.
Etymology. The specific epithet honors our friend, arachnologist Dr Regiane Saturnino, who collected the holotype. Noun in genitive case. Natural history. The single specimen was collected with pitfall trap in primary upland Amazonian Rain forest (Terra Firme) from Bagre, municipality in eastern Amazon, state of Pará, Brazil. Surazomus saturninoae sp. nov. is the third species of the genus collected from eastern Amazon. The two others were also collected from state of Pará, Brazil, S. paitit from the upland Amazonian Rain forest of Caxiuanã, and S. algodoal from the dry forest (Restinga) of Algodoal Island. Gustavo R. S. Ruiz and Roberta M. Valente. 2019. Description of A New Species of Surazomus (Arachnida: Schizomida), with Comments on Homology of Male Flagellum and Mating March Anchorage in the Genus. PLoS ONE 14(3): e0213268. DOI: 10.1371/journal.pone.0213268 | ||||||||||||||
| 11:55a | [Botany • 2019] Notes on the Genus Argostemma (Rubiaceae) from Lao PDR
ABSTRACT Three species of Argostemma (Rubiaceae) are newly recorded for Lao PDR: A. ebracteolatum, A. pictum and A. verticillatum. Descriptions and photographs of the new records are provided, and a key to Argostemma in Lao PDR is presented. KEYWORDS: Agrostemma, key, new record, Lao PDR, taxonomy INTRODUCTION Argostemma Wall., a genus of Rubiaceae of ca 100 species in the Old World tropics, is largely confined to the South-East Asia (Robbrecht, 1988) but with two species in tropical West Africa (Sridith & Puff, 2000; Mabberley, 2008). For Lao PDR, Newman et al. (2007a) and Newman et al. (2007b) first recorded a species of the genus, A. laeve Benn., from Khammouan province. Lanorsavanh & Chantaranothai (2013, 2016) recorded three species A. laotica Lanors. & Chantar., A. neurocalyx Miq. and A. siamense Puff from Bolikhamxai province, and, most recently, Tagane et al. (2017) recorded an additional species, A. lobbii Hook.f., from Bolikhamxai province. During botanical surveys of the first author in northern Lao PDR, Luangphrabang Province in July 2014 with S. Souvanhnakhoummane, and Luangnamtha Province in August 2015 with C. Phongoudome and in the southern Attapeu Province in June 2014 with V. Lamxay, many unnamed specimens were collectedand identified and include new records for Lao PDR, viz Argostemma ebracteolatum E.T.Geddes, A. pictum Wall. and A. verticillatum Wall. In 1999, Sridith indicated the type specimens of A. ebracteolatum, A. pictum, A. pubescens, A. repens and A. rotundifolium were holotypes, but this is an error to be corrected to lectotypes, nevertheless the lectotypification was effectively done by him.  NEW RECORDS: 1. Argostemma ebracteolatum E.T.Geddes, Bull. Misc. Inform. Kew 1927(4): 165. 1927. Type: Thailand, Chiang Mai, Mae Chaem (Me Cham), 14 Jul. 1922, Kerr 6276A (lectotype K! [K000760179], isolectotypes BK! [257307], BM! [BM000028695], designated by Sridith, 1999). Ecology.— On limestone wet rock in mixed deciduous, deciduous and dry evergreen forests, 540–1,100 m alt.; flowering July and fruiting July to August. Notes.— Argostemma ebracteolatum resembles A. thaithongae Sridith, endemic to Thailand, in the bell-shaped corolla and 5-merous flower, but differs in having not only being densely hairy on stem, leaves and inflorescence but also the exterior of the corolla. It is unlikely to be confused with other species in Laos as only A. neurocalyx has a bell-shaped corolla but is 4-merous. 2. Argostemma pictum Wall. in Roxb., Fl. Ind. (ed. Carey & Wall.) 2: 327. 1824. Type: Malaysia, Penang, 1892, Wallich, Numer. List 8392 (lectotypeK! [K000172892]; isolectotypes K! [K000172894], K-W! [K-W001125373] designated by Sridith, 1999), non Korth., 1851. Ecology.— On moist sandstone rocks with soil or on wet ground in evergreen forests, 100–410 m alt.; flowering June to July and fruiting June to August. Notes.— Argostemma pictum resembles A. neurocalyx but it distinguished by the wheel-shaped corolla which is 5-merous, and very strongly recurved with the tip coiled. 3. Argostemma verticillatum Wall. in Roxb., Fl. Ind. (ed. Carey & Wall.), 2: 325. 1824; Type: Nepal, Moreko, July 1821, Wallich, Numer. List 8394A(holotype K-W! [K-W001125376]). Fig. 2. Ecology.— On moist rocks near small water-fall in evergreen forests, 740–2,200 m alt., flowering and fruiting July to August. Notes.— Argostemma verticillatum is recognized by its curved filaments which its fused around the middle and forming a short filament tube. Because the Chinese species have free filaments (Chen & Taylor, 2011), therefore, we exclude China from the distribution information; further study is needed to clarify this character.
Soulivanh Lanorsavanh and Pranom Chantaranothai. 2019. Notes on the Genus Argostemma (Rubiaceae) from Lao PDR. Thai Forest Bulletin (Botany). 47(1); 29-33. DOI: 10.20531/tfb.2019.47.1.06   | ||||||||||||||
| 12:00p | [Paleontology • 2019] Iberodactylus andreui • A New Crested Pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the Radiation of the Clade Anhangueria
Abstract The pterosaur record from the Iberian Peninsula is mostly scarce and undefined, but in the last few years some new taxa have been described from different Lower Cretaceous sites of Spain. Here we describe a new genus and species of toothed pterodactyloid pterosaur from the Barremian of the Iberian Peninsula, Iberodactylus andreui gen. et sp. nov., that shows a close and rather unexpected relationship with Hamipterus tianshanensis from China. A review of the phylogenetic relationships of the Anhangueria reveals a new family of pterodactyloid pterosaurs, the Hamipteridae fam. nov. being recovered as sister-group of the Anhangueridae. This latter clade can be in turn divided into the new clades Anhanguerinae and Coloborhynchinae. The close relationships of Iberodactylus and Hamipterus shows an interesting palaeobiogeographical correlation between the Chinese and Iberian pterosaur faunas during the Barremian (Lower Cretaceous). The discovery of Iberodactylus strongly suggests that the clade Anhangueria has clear ancestral ties in eastern Laurasia.
Pterosauria Kaup, 1834. Pterodactyloidea Plieninger, 1901. Ornithocheiroidea Seeley, 1870 sensu Kellner (2003). Pteranodontoidea Marsh, 1876 sensu Kellner (2003). Lanceodontia Andres et al. (2014). Ornithocheirae Seeley, 1870 sensu Andres et al. (2014). Anhangueria Rodrigues & Kellner, 2013. Hamipteridae fam. nov. Branch-based definition: The most inclusive clade containing Hamipterus tianshanensis, but not Ludodactylus sibbicki, Coloborhynchus clavirostris, and Anhanguera blittersdorffi. Diagnosis: Crested anhanguerian pterodactyloids with the following synapomorphies: well-defined parallel and forward curved striae and sulci on the anterior region of the premaxillary crest, and an anterior rounded expansion of the anterior margin of the premaxillary crest. Included species: Hamipterus tianshanensis and Iberodactylus andreui gen. et sp. nov. Iberodactylus andreui gen. et sp. nov. Etymology: From the Iberian Peninsula and the Iberian System, where the specimen was recovered, and ‘dactylos’ (δάκτυλος), finger (ancient Greek), a common suffix in pterosaur names; in honour of Mr. Javier Andreu, a local collector who found the fossil. Holotype: Museo de Ciencias Naturales de la Universidad de Zaragoza (MPZ, Zaragoza, Spain) MPZ-2014/1; an anterior portion of a rostrum, including premaxillae –with a premaxillary crest– and maxillae, both with alveoli and broken teeth. Horizon and locality: Los Quiñones site, Morenillo limestones of the Blesa Formation, Barremian (Lower Cretaceous), Oliete sub-basin, Iberian Basin. Obón, Teruel Province, Aragón, northeast Spain. Diagnosis: Hamipterid pterodactyloid with the following autapomorphies: relatively deep premaxillary tip, premaxillary crest with its anterior margin curvature at an angle of about 80°.
Anhanguerinae clade nov. Stem-based definition: The most inclusive clade containing Anhanguera blittersdorffi but not Coloborhynchus clavirostris. Diagnosis: Anhanguerids with an enlarged fourth premaxillary tooth, larger than the fifth and sixth teeth and as large as or larger than the third tooth. Content: Anhanguera, Caulkicephalus, Cearadactylus, Guidraco, Liaoningopterus, Ludodactylus and Maaradactylus. Coloborhynchinae clade nov. Stem-based definition: The most inclusive clade containing Coloborhynchus clavirostris but not Anhanguera blittersdorffi or Ludodactylus sibbicki. Diagnosis: Anhanguerids with a quadrangular expansion of the premaxillary tip and a flat anterior surface of the rostrum24. Content: Coloborhynchus, Siroccopteryx and Uktenadactylus.
Borja Holgado, Rodrigo V. Pêgas, José Ignacio Canudo, Josep Fortuny, Taissa Rodrigues, Julio Company and Alexander W. A. Kellner. 2019. On A New Crested Pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the Radiation of the Clade Anhangueria. Scientific Reports. volume 9, 4940. DOI: 10.1038/s41598-019-41280-4   |














| << Previous Day |
2019/03/21 [Calendar] |
Next Day >> |