[Most Recent Entries] [Calendar View]
Wednesday, June 10th, 2020
| Time | Event | ||||||||||||
| 12:24a | [Botany • 2020] Winitia thailandana • Generic Status of Winitia (Annonaceae, Miliuseae) Reaffirmed by Molecular Phylogenetic Analysis, Including A New Species and A New Combination from Thailand
Abstract The generic status of Winitia Chaowasku (Annonaceae Juss., Miliuseae Hook.f. & Thomson) is reaffirmed by an extensive phylogenetic reconstruction using seven plastome regions (matK, ndhF, rbcL, ycf1 exons; trnL intron; psbA-trnH, trnL-trnF intergenic spacers) and including, among others, seven accessions of Winitia plus two accessions of its sister group, Stelechocarpus Hook.f. & Thomson. The results disclosed a maximally supported clade of Winitia, as well as of Stelechocarpus. The sister relationship of the two genera is still rather poorly supported and the branch uniting them is very short, whereas the branches leading to Winitia and to Stelechocarpus are relatively long, corresponding to their considerable morphological differences. Additionally, in Miliuseae there is a particular indel of eight continuous base pairs in the trnL-trnF intergenic spacer potentially diagnostic for generic discrimination, i.e., members in the same genus possess the same indel structure (absence or presence of a gap), and this indel differentiates Winitia from Stelechocarpus. Winitia cauliflora (Scheff.) Chaowasku appears polyphyletic and Winitia longipes (Craib) Chaowasku & Aongyong comb. nov. based on Stelechocarpus longipes Craib is consequently proposed. Furthermore, our phylogenetic data support a new species, Winitia thailandana Chaowasku & Aongyong sp. nov. from southern Thailand, which is described and illustrated. A key to genera in the sageraeoid clade (Sageraea-Winitia-Stelechocarpus) and a key to the four species of Winitia are provided. Keywords: identification key; Malmeoideae; phylogenetics; systematics; taxonomy Taxonomy Order Magnoliales Juss. ex Bercht. & J.Presl Family Annonaceae Juss., nom. cons. Tribe Miliuseae Hook.f. & Thomson Genus Winitia Chaowasku Winitia longipes (Craib) Chaowasku & Aongyong comb. nov. ≡ Stelechocarpus longipes Craib, The Journal of the Natural History Society of Siam 6: 43 (Craib 1923). Type: THAILAND · Khao Rum [Nakhon Si Thammarat Prov.]; fl; Native Collector s.n. (Herb. Eryl Smith 643); holotype: K[K000382080]; isotype: BK.

Winitia thailandana Chaowasku & Aongyong sp. nov. Diagnosis:Morphologically similar to Winitia expansa, differs by having a non-glaucous appearance on the petal adaxial surface (glaucous in W. expansa) and shallow pits on the adaxial surface of the inner petals (no pitted structures in W. expansa). Moreover, its inflorescences are mostly clustered on large roots and at the unswollen base of trunks, whereas the inflorescences of W. expansa are mostly clustered at the swollen base of trunks. Etymology: Named after Thailand, a country where this species is endemic.   Tanawat Chaowasku, Kithisak Aongyong, Anissara Damthongdee, Hathaichanok Jongsook and David M. Johnson. 2020. Generic Status of Winitia (Annonaceae, Miliuseae) Reaffirmed by Molecular Phylogenetic Analysis, Including A New Species and A New Combination from Thailand. European Journal of Taxonomy. 659; 1–23. DOI: 10.5852/ejt.2020.659   แสดทักษิณา Winitia thailandana Chaowasku & Aongyong พืชชนิดใหม่ของโลก พบที่เขาหินปูนจังหวัดสุราษฎร์ธานีเท่านั้ | ||||||||||||
| 1:33a | [Crustacea • 2020] On the Taxonomy of the Endemic Inle Lake Crab, Inlethelphusa acanthica (Kemp, 1918) (Brachyura: Potamidae) of Myanmar
Abstract The potamid freshwater crab, Inlethelphusa acanthica (Kemp, 1918), is endemic to Inle Lake, in Shan state, Myanmar, and was hitherto known only from the types and several small specimens collected in the 1930s. The species, the type of Inlethelphusa Yeo & Ng, 2007, is redescribed from a good series of fresh specimens. The taxonomy of the species is discussed and variations in the armature of the carapace anterolateral margin, male pleon, chela, as well as the male first gonopod structure are documented. Significantly, the male first gonopod of smaller specimens has the terminal segment straight and more cylindrical, with the tip truncate or weakly bifurcated, in contrast to that of larger males, which is more elongate, with the distal part gently curved upwards and the tip sharp. Key words: Decapoda, freshwater crab, redescription, variation, diagnostic characters, wetland conservation
 TAXONOMY Family Potamidae Ortmann, 1896 Subfamily Potamiscinae Bott, 1970 sensu Yeo & Ng, 2004 Inlethelphusa Yeo & Ng, 2007 Type species. Potamon (Potamon) acanthicum Kemp, 1918, by monotypy. Diagnosis. Carapace broader than long, relatively low; dorsal surface gently convex; epigastric cristae low, distinct, not sharp, anterior to postorbital cristae, distinctly separated from postorbital cristae by groove; postorbital cristae low, blunt, not confluent with epibranchial teeth; external orbital angle well developed, separate from rest of anterolateral margin by deep cleft; epibranchial tooth well developed, spiniform; anterolateral margins prominently serrated, with anterior serrations larger; antennular fossae subrectangular, broad; epistome posterior margin with low but distinct median tooth; endostomial ridge absent. Third maxilliped with long exopod, with long flagellum. Ambulatory legs short; merus without subdistal spine on dorsal margin. Suture between male thoracic sternites 2 and 3 distinct; thoracic sternite 8 completely separated by a longitudinal medial line; sternopleonal cavity almost reaching imaginary line joining anterior edges of cheliped bases; male pleon triangular. G1 subterminal segment with distal third bent outward, appears neck-like, being distinctly tapered or constricted; G1 terminal segment slender, elongate, tubular to tapering, tip truncate or sharp, upcurved, dorsal flap absent. G2 longer than G1, distal segment distinctly less than half length of basal segment. Inlethelphusa acanthica (Kemp, 1918) (Figs. 1–8) Potamon (Potamon) acanthicum Kemp, 1918: 85, fig. 2, pl. 24 figs. 3, 4. Lobothelphusa acanthia – Bott, 1970: 150; Yeo & Ng, 1999: 640. Inlethelphusa acanthica – Yeo & Ng, 2007: 283, fig. 6; Ng et al., 2008: 163. Ecological notes. Kemp (1918: 89) notes that this “is apparently the only crab that makes its way into the central region of the Inle Lake, on the bottom of which it is occasionally found. It is more abundant among the roots of the floating islands at the edge of the lake, and also frequents small hill-streams.” The series of recent specimens (ZRC 2018.1370) were from the centre of the lake and were collected in the nets of fishermen. Two other specimens (ZRC 2018.0736) were from small streams flowing into Inle Lake. Nothing else is known about its ecology. This potamid species occupies a lacustrine niche in northern Indochina that, in other parts of Southeast Asia, is normally filled by crabs of the family Gecarcinucidae Rathbun, 1904. The chelae of I. acanthica are gaped in large males and females (Fig. 3D, E), similar to those of the analagous lentic gecarcinucids found in swamps and lakes in Thailand and Cambodia, and are not specifically adapted for feeding on molluscs (see Ng & Naiyanetr, 1993; Ng, 1995). In ancient lakes in Sulawesi, molluscivorous species have large molariform teeth at the base of the gaping fingers (see Chia & Ng, 2006; Schubart & Ng, 2008), while there is no evidence of incipient development or evolution of any such molariform teeth in I. acanthica.  Peter K. L. Ng, Win Mar and Darren C. J. Yeo. 2020. On the Taxonomy of the Endemic Inle Lake Crab, Inlethelphusa acanthica (Kemp, 1918) (Crustacea: Brachyura: Potamidae) of Myanmar. RAFFLES BULLETIN OF ZOOLOGY. 68; 453–463.   | ||||||||||||
| 1:34a | [Paleontology • 2020] Dietary Palaeoecology of An Early Cretaceous Armoured Dinosaur (Ornithischia; Nodosauridae) Based on Floral Analysis of Stomach Contents
Abstract The exceptionally well-preserved holotype of the armoured dinosaur Borealopelta markmitchelli (Ornithischia; Nodosauridae) from the Early Cretaceous (Clearwater Formation) of northern Alberta preserves a distinct mass within the abdominal cavity. Fourteen independent criteria (including: co-allochthony, anatomical position, gastroliths) support the interpretation of this mass as ingested stomach contents—a cololite. Palynomorphs in the cololite are a subset of the more diverse external sample. Analysis of the cololite documents well-preserved plant material dominated by leaf tissue (88%), including intact sporangia, leaf cross-sections and cuticle, but also including stems, wood and charcoal. The leaf fraction is dominated (85%) by leptosporangiate ferns (subclass Polypodiidae), with low cycad–cycadophyte (3%) and trace conifer foliage. These data represent the most well-supported and detailed direct evidence of diet in an herbivorous dinosaur. Details of the dietary palaeoecology of this nodosaur are revealed, including: selective feeding on ferns; preferential ingestion of leptosporangiate ferns to the exclusion of Osmundaceae and eusporangiate ferns such as Marattiaceae; and incidental consumption of cycad–cycadophyte and conifer leaves. The presence of significant (6%) charcoal may represent the dietary use of recently burned conifer forest undergoing fern succession, early evidence of a fire succession ecology, as is associated with many modern large herbivores. Keywords: cololite, Ankylosauria, diet, Canada, Clearwater Formation, Cretaceous
Caleb M. Brown, David R. Greenwood, Jessica E. Kalyniuk, Dennis R. Braman, Donald M. Henderson, Cathy L. Greenwood and James F. Basinger. 2020. Dietary Palaeoecology of An Early Cretaceous Armoured Dinosaur (Ornithischia; Nodosauridae) Based on Floral Analysis of Stomach Contents. Royal Society Open Science. DOI: 10.1098/rsos.200305 A Nodosaur’s Last Meal The world’s best-preserved armoured dinosaur, the nodosaur Borealopelta markmitchelli, continues to answer important questions about its biology and behaviour. | ||||||||||||
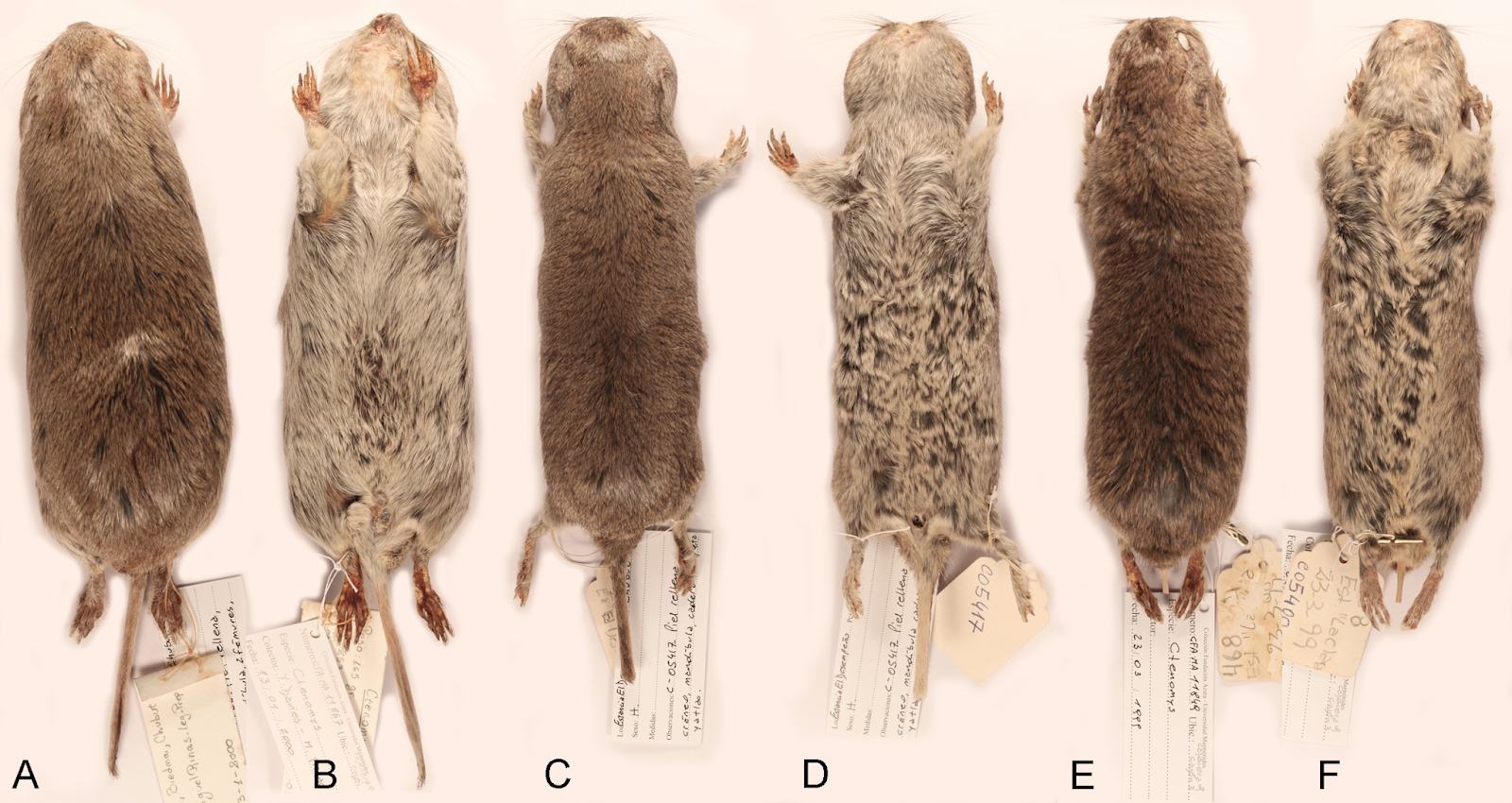
| 2:00a | [Mammalogy • 2020] Uncovering the Species Diversity of Subterranean Rodents at the End of the World: Three New Species of Patagonian Tuco-tucos (Rodentia, Hystricomorpha, Ctenomys)
Abstract Ctenomys Blainville 1826 is one of the most diverse genera of South American caviomorph rodents. Currently, six species of this genus are reported from Patagonia, south of 42°S. In this contribution, we assessed the taxonomic status of several populations from eastern and central Chubut province, northern Patagonia. Based on phylogenetic analyses of DNA sequences, morphology assessment (qualitative and quantitative), and previously published karyological data, we describe three new species of this genus, one formed by two subspecies, endemic to northern Patagonia. In addition, we include C. coyhaiquensis Kelt and Gallardo 1994 into the synonymy of C. sericeus J.A. Allen 1903. Finally, we discussed the need for additional integrative approaches, including field collection of specimens, to better understand the diversity of this highly speciose rodent genus.
Ctenomys bidaui n. sp. Bidau’s tuco-tuco Tuco-Tuco de Bidau Morphological diagnosis—A medium-sized tuco-tuco of the C. magellanicus species group with moderately differentiated dorsal and ventral colorations; dorsum Light Brownish Olive to Brownish Olive; venter Pale Olive Buff with Gray colored basal hairs. Skull strongly built; interorbital processes of frontals slightly developed; zygomatic arches robust; premaxillo-frontal suture at the level of the naso-frontal suture; interparietal broad and short; incisive foramina moderately short and broad, recessed in a common fossa of straight outer borders and incompletely separated by a bony septum; interpremaxillary foramen large; auditory bullae inflated and ovate. Distribution—Known only from three localities near coastal areas of Península de Valdés, Chubut, Argentina (Fig. 1). Possibly, also correspond to this species the late Holocene fossil remains referred by Udrizar Sauthier & D’Agostino (2017) from this same general area. Etymology—We named this species in honor of the late Claudio J. Bidau (1953-2018), an Argentinian biologist with an extensive and varied scientific production, of which an important fraction is aimed to elucidate the complex evolutionary history of the genus Ctenomys. Claudio was a much-appreciated member of the South American community of mammalogists where he is well remembered. The species name is a patronym in the genitive singular. Ctenomys contrerasi n. sp. Contreras’tuco-tuco Tuco-Tuco de Contreras Morphological diagnosis—A small to medium sized tuco-tuco of the C. magellanicus species group with dorsal and ventral colorations moderately differentiated; dorsum Brownish Olive to Olive or Tawny Olive; venter Pale Olive Buff with Gray colored basal hairs. Skull moderately robust, interorbital region with posteriorly divergent outer margins; premaxillo-frontal suture placed slightly to well behind from the naso-frontal suture; zygomatic arch thin to moderately robust, with slightly developed postorbital and mandibular processes of jugal and a conspicuous zygomatic depression; interparietal completely fused; incisive foramina moderately long and narrow, recessed in a common fossa of straight to slightly convex outer borders and completely separated by a thin bony septum; interpremaxillary foramen small to large; paraoccipital process fan-shaped; auditory bullae inflated, and pyrifom. Distribution—This species has an apparently disjunct distribution, being recorded at four localities close to the Atlantic coast, between the Chubut river in the south and the Ameghino Isthmus to the north, and other two populations in west-central Chubut, south of the Chubut river (Fig. 1). Both distributional areas are about 335 km apart. Etymology—This species of Ctenomys is named in honor of Julio R. Contreras (1933-2017), an Argentinean mammalogist and ornithologist who dedicated more than 45 years of his life to the study of the taxonomy, systematics, and biogeography of the genus Ctenomys (see Teta & Ríos, 2019). Contreras described more than a dozen of new species of tuco-tucos, both from Argentina and Paraguay. Together with C. Bidau (Contreras & Bidau, 1999), he authored one of the first attempts to summarize the complex evolutionary history of this genus, proposing a general hypothesis about its diversification. The species name is a patronym in the genitive singular. Ctenomys contrerasi contrerasi n. subsp. Ctenomys contrerasi navonae n. subsp. Etymology—This subspecies of Ctenomys is named in honor of our colleague and friend Graciela T. Navone, an Argentinean parasitologist with a large career studying small mammal endoparasites. Graciela is also a prominent and active member of the Sociedad Argentina para el Estudio de los Mamíferos (SAREM). The species name is a patronym in the genitive singular. Ctenomys thalesi n. sp. Thales’s tuco-tuco Tuco-Tuco de Thales Morphological diagnosis— A small-sized tuco-tuco of the C. magellanicus species group with dorsal and ventral coloration moderately differentiated; dorsum Light Brownish Olive; venter Pale Olive Buff with gray colored basal hairs. Skull moderately robust, interorbital region with posteriorly divergent outer margins; premaxillo-frontal suture placed slightly behind the naso-frontal suture; zygomatic arch thin, with slightly developed postorbital and mandibular processes of jugal and a conspicuous zygomatic depression; interparietal completely fused; incisive foramina moderately short and narrow, recessed in a common fossa of nearly convex outer borders and completely separated by a thin bony septum; interpremaxillary foramen small to absent; paraoccipital process hook-shaped; auditory bullae inflated, and pyrifom. Distribution— Known only from two localities on northeastern Chubut province, close to the Atlantic coast, south of Chubut river (Fig. 1). Etymology— We name this species in honor of Thales Renato Ochotorena de Freitas, a Brazilian geneticist who leads a productive research program mostly centered on Brazilian species of Ctenomys, covering among others, aspects of taxonomy, cytogenetics, speciation, phylogeography, and conservation genetics. The species name is a patronym in the genitive singular. Ctenomys sericeus Allen, 1903 Silky tuco-tuco Tuco-Tuco sedoso Morphological diagnosis—pelage short, soft, and glossy (Fig. 5S, on Data S2); general color above Olive Brown to Sepia strongly varied with Black, the hairs being Dark Gray for the basal three fourths, then banded narrowly with pale Yellowish Brown, and tipped with Black; top of nose and top of head like median dorsal region, which is darker than the sides, sometimes forming a dark median dorsal band extending from the nose to the base of the tail; flanks lighter than dorsum and venter Isabella; ears very small, blackish; upper surface of feet grayish to yellowish; tail Tawny Olive, with a median dusky stripe along the apical half of the upper surface. Skull moderately robust (Fig. 6S, on Data S2), interorbital region with posteriorly divergent outer margins; premaxillo-frontal suture placed behind from the naso-frontal suture; zygomatic arches robust, with conspicuously and moderately developed postorbital and mandibular processes of jugal, respectively, and a well-marked zygomatic depression; interparietal absent to very small; incisive foramina moderately short and broad, recessed in a common fossa of convex outer borders and completely separated by a thin bony septum; interpremaxillary foramen large to inconspicuous; paraoccipital hook-shaped; auditory bullae inflated, and pyrifom. Distribution— C. sericeus occurs in open shrubby to herbaceous steppes from southwestern Chubut (Argentina) in the north to the northern margin of the Santa Cruz river (Santa Cruz, Argentina) in the south, and adjacent open areas of Aysen, Chile (Fig. 1). Conclusions: The integrative analyses of morphological, molecular, and karyotipic data of Patagonian specimens of Ctenomys allowed as to describe three new species endemics to the open areas of northern Patagonia. The three new species belong to the C. magellanicus species group. In addition, we consider the geographically restricted C. coyhaiquensis (Kelt & Gallardo, 1994) as a junior synonym of the widespread C. sericeus (Allen, 1903). Our results also shown that as currently understood, C. haigi is likely a composite of two lineages of species level; tentatively, we refer to then as C. haigi s.s. and C. cf. C. lentulus. Our findings, together with the fact that large Patagonian areas still remain unstudied, suggest that the diversity of Patagonian species of Ctenomys is only partially understood. Therefore, to fill in this gap of knowledge, it is needed to carry out additional integrative taxonomic studies, based on the field collection of additional specimens. Pablo Teta and Guillermo D’Elía. 2020. Uncovering the Species Diversity of Subterranean Rodents at the End of the World: Three New Species of Patagonian Tuco-tucos (Rodentia, Hystricomorpha, Ctenomys). PeerJ. 8:e9259. DOI: 10.7717/peerj.9259 | ||||||||||||
| 2:02a | [Botany • 2020] Tillandsia dichromantha (Tillandsioideae; Bromeliaceae) • A New Species from the State of Oaxaca, Mexico
Abstract Tillandsia dichromantha, a new species from the state of Oaxaca, Mexico, is described and illustrated. The proposed taxon is compared with T. achyrostachys and T. ilseana, species with which it has some similarities. Images and a distribution map are included. Keywords: Endemic, Monocots, Poales
  Tillandsia dichromantha Hern.-Cárdenas, López-Ferr. & Espejo, sp. nov. The new species is similar to Tillandsia ilseana and T. achyrostachys but differs in the height of the flowering plants (45–55 vs. ± 100, and 24–45 cm), the number of spikes (6–9 vs. (4)9–15, and one), the ecarinate and conspicuosly nerved floral bracts (vs. carinatesmooth, and ecarinate-conspicuosly nerved), the floral bracts width (1.3–1.6 vs. 0.7–0.8, and 1.6–2.3 cm), the sepals size (2.3–2.5 × 0.5–0.6 vs. ±2.8 × 0.8–0.9, and 2.7–2.9 × 0.6–0.7 cm), the petal color (violet + green-chartreuse vs. very pale violet [T. ilseana], and chartreuse green [T. achyrostachys]), and in the style length (6–6.5 vs. 5.5–6, and 4.5–5 cm).
Distribution and habitat:— Tillandsia dichromantha is only known from Oaxaca (Figure 2): the Sierra Madre del Sur in the municipalities of San Juan Mixtepec; and Santos Reyes Tepejillo, in the district of Juxtlahuaca, where it grows epiphytically on Quercus spp. and Acacia sp. in dry oak (Quercus) forests, Pinus forests, and tropical deciduous forests with species of Arecaceae, Ipomoea species, Bursera species, and Juniperus species (Figures 1A–B). Tillandsia dichromantha grows among other Tillandsia species like T. achyrostachys, T. dugesii Baker (1887: 278), T. recurvata L. (1762: 410), and T. usneoides L. (1762: 411). Plants of T. dichromantha inhabit localities between 1,730 and 1,960 m elevation and blooms from December to January. It should be mentioned that we found only three individuals during all the visits to the zone along a year, so probably the plants are scarce. Etymology:— The specific epithet refers to the bicolorous petals, violet and chartreuse green, that distinguish this new species. Rodrigo Alejandro Hernández-Cárdenas, Adolfo Espejo-Serna, Ana Rosa López-Ferrari and Sofía Ana Lucrecia Lara-Godínez. 2020. Tillandsia dichromantha (Tillandsioideae; Bromeliaceae), A New Species from the State of Oaxaca, Mexico. Phytotaxa. 447(2); 81–87. DOI: 10.11646/phytotaxa.447.2.1   | ||||||||||||
| 2:03a | [Herpetology • 2020] Platypelis laetus • Frogs of the Genus Platypelis from the Sorata Massif in northern Madagascar: Description of A New Species and Reports of Range Extensions
Abstract We describe a new species of arboreal microhylid frog, genus Platypelis, from northeastern Madagascar and report the expansion of distribution ranges of two other species. Platypelis laetus sp. nov. is small to medium-sized (24.3–25.6 mm snout-vent length) compared to other Platypelis, exhibits a greenish colored throat and was found in bamboo forest of the Sorata Massif. Its advertisement call consists of a single short tonal note repeated at regular intervals in long call series. Based on DNA sequences of a fragment of the mitochondrial 16S rRNA gene, the new species was placed in a clade with Platypelis olgae from the Tsaratanana Massif, and with two other, unconfirmed candidate species from the Sorata Massif and from Andravory, herein named Platypelis sp. Ca12 and Ca13. Molecular divergences among these lineages were substantial, amounting to 7.6‒8.1% uncorrected 16S p-distance to the closest nominal species, P. olgae, from which the new species is also distinguished by a lack of allele sharing in the nuclear RAG-1 gene. We also provide new records of Platypelis alticola and P. tsaratananaensis from the Sorata Massif, supported by molecular analysis. This confirms a wider distribution of these two species that previously were considered to be endemic to the Tsaratanana Massif. However, their populations in Sorata were characterized by a certain degree of genetic differentiation from Tsaratanana populations suggesting they require more detailed taxonomic assessment. Key Words: Amphibia, Anura, Cophylinae, distribution, Microhylidae, molecular genetics, Platypelis laetus sp. nov., Platypelis alticola, Platypelis tsaratananaensis, Sorata, systematics 
Platypelis laetus sp. nov. Diagnosis: Assigned to the genus Platypelis in the microhylid subfamily Cophylinae based on enlarged terminal discs on fingers and toes, absence of nuptial pads, and molecular phylogenetic relationships. The species can be identified among other cophylines by the combination of the following character states: (1) medium-sized species (adult male SVL 24.3–25.6 mm); (2) manus with second finger slightly shorter than fourth and pes with third toe much shorter to very slightly shorter than fifth; (3) males with prepollical tubercle but lacking a finger-like prepollex as typical for Anodonthyla; (4) throat greenish in life; (5) chest and anterior belly translucent gray, with distinct white spotting that is absent on the posterior belly; (5) absence of red color on limbs and ventral side. Distribution: The species is known only from the Sorata Massif, northern Madagascar, at elevations of 1339‒1541 m above sea level (Fig. 2). Natural history: Platypelis laetus sp. nov. occurs in rainforest on the Sorata Massif, but most of the specimens were collected in the bamboo forest of the massif. The holotype was calling from a bamboo hole at about 5 m above the ground. The bamboo node was occupied by another specimen, was water-filled, and contained 35 whitish eggs and embryos, probably of this species, in at least two different developmental stages (Fig. 3h). Another male was found on a palm tree at around 4 m above the ground. Calling occurred around dusk. Etymology: The specific epithet is a masculine Latin adjective meaning “happy”. The new species is so named in reflection of the joy and happiness of the first author to get to work on the cophyline microhylid frogs of Madagascar. Andolalao Rakotoarison, Mark D. Scherz, Jörn Köhler, Fanomezana M. Ratsoavina, Oliver Hawlitschek, Steven Megson, Miguel Vences and Frank Glaw. 2020. Frogs of the Genus Platypelis from the Sorata Massif in northern Madagascar: Description of A New Species and Reports of Range Extensions. Zoosystematics and Evolution. 96(1): 263-274. DOI: 10.3897/zse.96.47088  | ||||||||||||
| 2:04a | [Botany • 2020] Michaelmoelleria vietnamensis (Gesneriaceae) • A New Lithophilous Dwelling Genus and Species with Zigzag Corolla Tube from southern Vietnam
Abstract Michaelmoelleria, a new genus from southern Vietnam is described with a single species, M. vietnamensis. The new genus is morphologically most similar to Deinostigma and Tribounia but it differs from the latter two by having four fertile stamens. Nuclear ribosomal internal transcribed spacer (ITS) region and plastid trnL-F intron spacer (trnL-F) DNA sequence data from the new genus and eighty-seven species representing 42 genera within tribe Didymocarpeae are used to resolve its generic placement. The molecular evidence reveals that it is most closely related to Cathayanthe rather than Deinostigma and Tribounia. The chromosome number is counted as 2n = 36 that further clarified its distinction comparing to the related genera within tribe Didymocarpeae. A global conservation assessment is also performed and classifies Michaelmoelleria vietnamensis as Critically Endangered (CR). Keywords: Cathayanthe, cliff-dwelling, Deinostigma, Didymocarpoideae, flora of Vietnam, IUCN, phylogeny, Tribounia

Michaelmoelleria F. Wen, Y.G. Wei & T.V. Do, gen. nov. Diagnosis: Michaelmoelleria resembles monotype genus Cathayanthe, but differs from the latter by leaf blade glabrous (vs. sericeous to pubescent in Cathayanthe, same as order followings); calyx actinomorphic (vs. zygomorphic); fertile stamens 4 (vs. 2), stigma 2, both developed ligulate (vs. 1, subcapitate, divided on 1 side); capsule long linear (vs. fleshly, narrowly ellipsoid). The new genus is also morphologically similar to Deinostigma and Tribounia, but is easily distinguished from both by having corolla tube narrowly curving to zigzag infundibuliform-tubular, and bent at about 90° angle in the middle of corolla tube (vs. infundibuliform in Deinostigma; of a narrow lower tube which widens into an infundibuliform & upper tube which has a prominent boss on the dorsal surface in Tribounia, same as order followings), fertile stamens number 4 (vs. 2; 2) and stigma 2-lobed, lobes often gathering together (vs. upper lip usually vestigial and only lower lip developing, broad, flat and weakly 2-lobed; capitate). Type and only known species: Michaelmoelleria vietnamensis F. Wen, Z.B. Xin & T.V. Do, sp. nov. Etymology: Michaelmoelleria was named in honor of Prof./Dr. Michael Möller from the Royal Botanic Garden Edinburgh. He is a well-known botanist studying Old World Gesneriaceae, especially in Africa (Madagascar) and Asia (China), and mentor of the senior author from the 1990s to the present. “Michaelmoeller-” (means “Michael Möller”) stands for his full name. “moeller” is the English modification of the German family name, “Möller”. Initially, we planned to use “Moelleria” as the genus name. However, this name was used in different places three times. They are Moelleria Cleve (Bacillariophyta, incertae sedis) [non Moelleria Scop. (Spermatophyta, Flacourtiaceae) (≡ Iroucana Aubl.)]; [nec Moelleria Bres. (Fungi, Clavicipitaceae) (≡ Moelleriella Bres.)] [nec Moelleria (Freng.) Freng. (Bacillariophyta, Naviculaceae)] (Blanco and Wetzel 2016). Thus, to prevent confusion with those mentioned above, three existing and existed “Moelleria”, we consider that using the variant of Dr. Michael Möller’s full name, “Michaelmoeller”, to name this new genus to be most appropriate. Distribution and habitat: Endemic to southern Vietnam, under broadleaved forests in a montane granite area at 140–200 m altitude. Michaelmoelleria vietnamensis F. Wen, Z.B. Xin & T.V. Do, sp. nov. Etymology: The genus is named for the famous botanist, Dr. / Prof. Michael Möller, from the Royal Botanic Garden Edinburgh, and the species is named for Vietnam, which holds the first discovered and only known location for the species.
  Fang Wen, Zi-Bing Xin, Long-Fei Fu, Shu Li, Lan-Ying Su, Stephen Maciejewski, Zhang-Jie Huang, Truong Van Do and Yi-Gang Wei. 2020. Michaelmoelleria (Gesneriaceae), A New Lithophilous Dwelling Genus and Species with Zigzag Corolla Tube from southern Vietnam. PhytoKeys. 146: 89-107. DOI: 10.3897/phytokeys.146.49731   | ||||||||||||
| 9:52a | [PaleoMammalogy • 2020] Juchuysillu arenalesensis • A New Typothere Notoungulate (Mammalia: Interatheriidae) from the Miocene Nazareno Formation of Southern Bolivia
Abstract We describe a new interatheriid notoungulate, Juchuysillu arenalesensis gen. et sp. nov., based on six partial upper and lower dentitions from the early to middle Miocene Nazareno Formation of southern Bolivia. A specimen is also referred to J. arenalesensis from the early middle Miocene (Langhian) locality of Cerdas, Bolivia (ca. 100 km to the northwest). The new species is distinguished by its very small size (ca. 15% smaller than Protypotherium minutum) and the unique combination of shallow ectoloph sulci on P3-4, length of M1 > M2 > M3, trapezoidal upper molars, and absence of a buccal talonid sulcus on m3. A second, larger interatheriid species is present at Nazareno but is not represented by remains sufficiently complete for a more precise identification. A phylogenetic analysis indicates that J. arenalesensis represents a unique lineage within Interatheriinae that diverged after Protypotherium sinclairi but before Miocochilius anomopodus, Caenophilus tripartitus, Miocochilius federicoi, and several species of Protypotherium, including Protypotherium australe. Juchuysillu arenalesensis is one of two small (< ca. 3 kg) typotheres at Nazareno; its body mass is estimated at 1.1 kg. An updated faunal list for the Nazareno Formation includes 19 species of mammals pertaining to seven orders and 15 families; fragmentary turtle (Testudinidae?) and bird (Phorusrhacidae) remains are also present. The precise age of Nazareno Formation vertebrates is unknown, but the presence of at least three notoungulate species shared with Cerdas suggests a similar (∼16–15 Ma) age for the fossil-bearing levels.     Darin A. Croft and Federico Anaya. 2020. A New Typothere Notoungulate (Mammalia: Interatheriidae), from the Miocene Nazareno Formation of Southern Bolivia. Ameghiniana. 57(2); 189-208. DOI: 10.5710/AMGH.11.01.2020.3271   |

























{kind=link}
| << Previous Day |
2020/06/10 [Calendar] |
Next Day >> |