[Most Recent Entries] [Calendar View]
Wednesday, April 28th, 2021
| Time | Event | ||||||||||
| 1:28a | [Herpetology • 2021] Pedioplanis branchi & P. mayeri • Lizards of A Different Stripe: Phylogenetics of the Pedioplanis undata Species Complex (Squamata, Lacertidae), with the Description of Two New Species
Abstract The lacertid genus Pedioplanis is a moderately speciose group of small-bodied, cryptically-colored lizards found in arid habitats throughout southern Africa. Previous phylogenetic work on Pedioplanis has determined its placement within the broader context of the Lacertidae, but interspecific relations within the genus remain unsettled, particularly within the P. undata species complex, a group largely endemic to Namibia. We greatly expanded taxon sampling for members of the P. undata complex and other Pedioplanis, and generated molecular sequence data from 1,937 bp of mtDNA (ND2 and cyt b) and 2,015 bp of nDNA (KIF24, PRLR, RAG-1) which were combined with sequences from GenBank resulting in a final dataset of 455 individuals. Both maximum likelihood and Bayesian analyses recover similar phylogenetic results and reveal the polyphyly of P. undata and P. inornata as presently construed. We confirm that P. husabensis is sister to the group comprising the P. undata complex plus the Angolan sister species P. huntleyi + P. haackei and demonstrate that P. benguelensis lies outside of this clade in its entirety. The complex itself comprises six species including P. undata, P. inornata, P. rubens, P. gaerdesi and two previously undescribed entities. Based on divergence date estimates, the P. undata species complex began diversifying in the late Miocene (5.3 ± 1.6 MYA) with the most recent cladogenetic events dating to the Pliocene (2.6 ± 1.0 MYA), making this assemblage relatively young compared to the genus Pedioplanis as a whole, the origin of which dates back to the mid-Miocene (13.5 ± 1.8 MYA). Using an integrative approach, we here describe Pedioplanis branchi sp. nov. and Pedioplanis mayeri sp. nov. representing northern populations previously assigned to P. inornata and P. undata, respectively. These entities were first flagged as possible new species by Berger-Dell’mour and Mayer over thirty years ago but were never formally described. The new species are supported chiefly by differences in coloration and by unique amino acid substitutions. We provide comprehensive maps depicting historical records based on museum specimens plus new records from this study for all members of the P. undata complex and P. husabensis. We suggest that climatic oscillations of the Upper Miocene and Pliocene-Pleistocene era in concert with the formation of biogeographic barriers have led to population isolation, gene flow restrictions and ultimately cladogenesis in the P. undata complex. Key Words: Biogeography, molecular phylogeny, phylogenetics, southern Africa, species description, taxonomy
Pedioplanis inornata “South”/P. inornata “Central” The type series of Eremias inornata Roux, 1907 comprises eight specimens collected from “Oranje-Fluß, Kl.-Namaqualand” [the Orange River, Little Namaqualand, Northern Cape, Province, South Africa]. Syntype ZMA11049 (Zoological Museum Amsterdam) was subsequently designated as the lectotype (Daan and Hillenius 1966). The ZMA collections have subsequently been incorporated into those of the Naturalis Biodiversity Center in Leiden (RMNH). The geographic origin of the specimens and the relatively detailed description and accompanying plate, clearly tie the name-bearing lectotype of Pedioplanis inornata to the P. inornata “South” clade. No other nomina are currently within the synonymy of Pedioplanis inornata and none are available for application to the P. inornata “Central” clade, which is here described as: Pedioplanis branchi sp. nov. Etymology: The specific epithet is a patronym formed in the genitive singular honoring our friend and colleague, the British-born South African herpetologist, William Roy Branch (1946–2018), in recognition of his many contributions to African herpetology and in remembrance of many happy trips in the field together. Distribution: Pedioplanis branchi sp. nov. is endemic to the Erongo Region in Namibia. Its range stretches from just south of the Swakop River in the South, where it occurs in parapatry with P. husabensis, through the pro-Namib, to the Ugab River and the Brandberg in the north and Mount Erongo and Otjimbingwe in the east, probably occurring in parapatry with P. mayeri all along its northeastern border (see Fig. 6 for map of locality records).
Pedioplanis undata “South”/P. undata “North” Mayer and Böhme (2000) examined the type material of Lacerta undata Smith, 1838 and noted that the rediscovered syntype specimens in the National Museums of Scotland collection were, in fact, representatives of the taxon long associated with the name Pedioplanis lineoocellata pulchella (Gray, 1845) and now, based on the findings of Edwards (2013), considered as P. lineoocellata (Duméril & Bibron, 1839). They proposed to conserve the current usage of names within Pedioplanis by calling on the ICZN in Case 3085 to set aside the original type designation and recognize the designation of a neotype for L. undata. Opinion 1992 of the ICZN (Anonymous 2002) accepted this solution and thus the neotype was fixed as Naturhistorisches Museum Wien (NMW) 31886 from “near Windhoek, Namibia.” This links the name P. undata to the P. undata “South” clade. The assignment of the name Pedioplanis undata to the more restricted, southern clade is notable in that the predominant concept of the species based on previous studies of P. undata is largely affiliated with populations and representative specimens from the more broadly distributed northern clade (Branch 1998; Makokha et al. 2007; Conradie et al. 2012). In these cases the presence of bold dorsal stripes was considered diagnostic for the species, however based on our results this character is now largely applicable to the unnamed northern clade, and is exhibited inconsistently and with variation among true P. undata. No other nomina are currently within the synonymy of Pedioplanis undata and none are available for application to the P. undata “North” clade, which we describe as: Pedioplanis mayeri sp. nov. Etymology: The specific epithet is a patronym formed in the genitive singular honoring our friend and colleague, the Austrian lacertid specialist Werner Mayer (1943–2015), who first recognized the distinctiveness of his namesake species and whose contributions to the study of Pedioplanis have been seminal. Distribution: Pedioplanis mayeri sp. nov. is endemic to northern Namibia and occurs from south of the Kunene River and east of the Namib Desert along the eastern side of the escarpment, thence throughout the eastern Kunene Region, entering the northeastern parts of the Erongo Region and east through the Otjozondjupa Region, reaching at least as far east as Oshikango (TM17028) in the north, Gobabis (Omaheke Region) in the south-east, and Nauchas in the south. It does not enter the Kalahari dune fields, and is possibly absent from the Khomas Hochland, where it is replaced by P. undata (see Fig. 6 for map of locality records).
  Jackie L. Childers, Sebastian Kirchhof and Aaron M. Bauer. 2021. Lizards of A Different Stripe: Phylogenetics of the Pedioplanis undata Species Complex (Squamata, Lacertidae), with the Description of Two New Species. Zoosystematics and Evolution. 97(1): 249-272. DOI: 10.3897/zse.97.61351  | ||||||||||
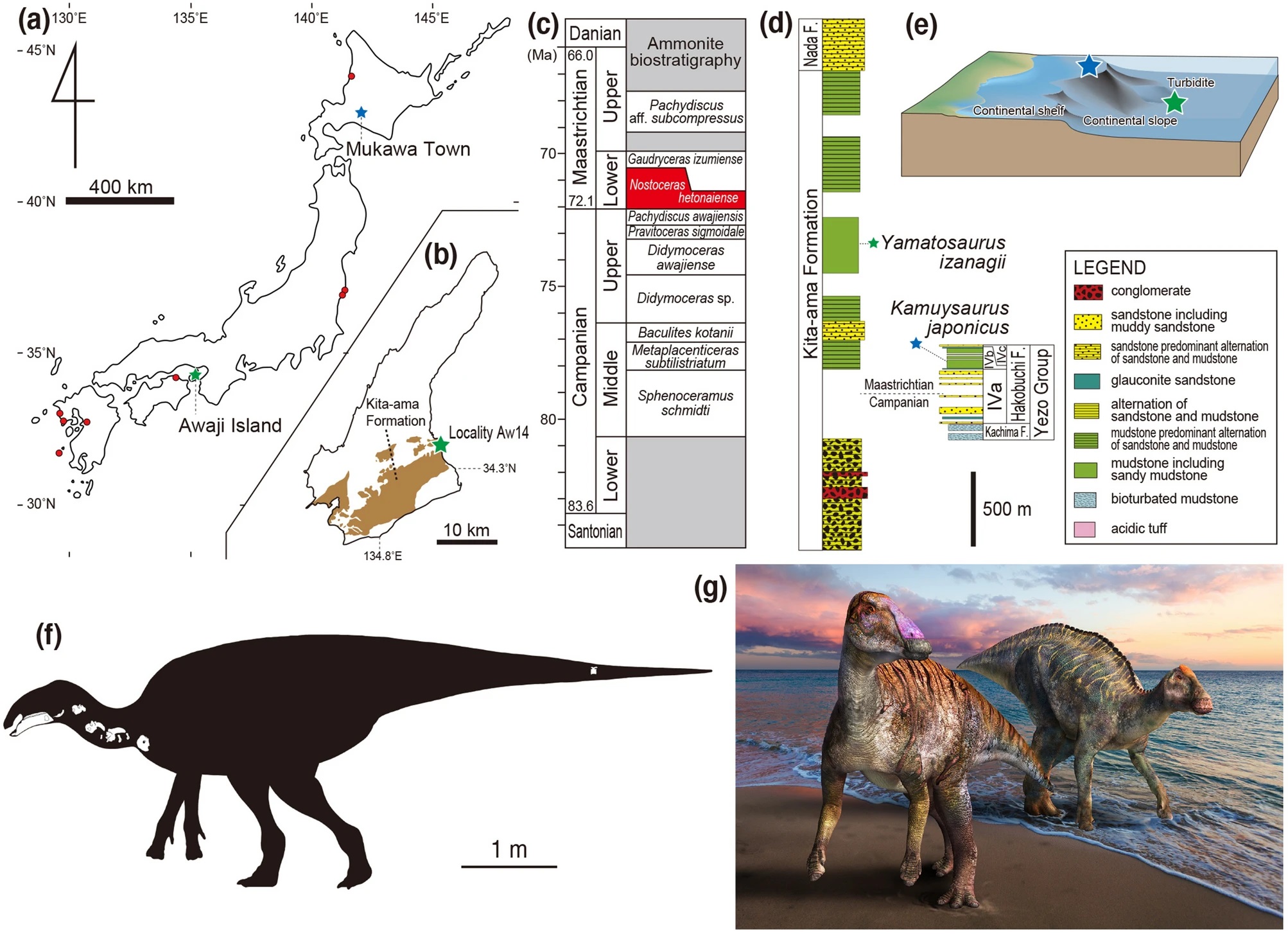
| 1:33a | [Paleontology • 2021] Yamatosaurus izanagii • A New Basal Hadrosaurid (Dinosauria: Ornithischia) from the latest Cretaceous Kita-ama Formation in Japan implies the Origin of Hadrosaurids
Abstract Here we describe a partial hadrosaurid skeleton from the marine Maastrichtian Kita-ama Formation in Japan as a new taxon, Yamatosaurus izanagii gen. et sp. nov., based on unique characters in the dentition. Our phylogenetic analysis demonstrates Yamatosaurus izanagii belongs to Hadrosauridae, composed of Hadrosaurus foulkii + (Yamatosaurus izanagii + (Saurolophinae + Lambeosaurin   
Systematic paleontology Dinosauria Owen, 1842 Ornithischia Seeley, 1888 Ornithopoda Marsh, 1881 Hadrosauridae Cope, 1869 Yamatosaurus izanagii gen. et sp. nov. Holotype: MNHAH D1-033516, a right dentary, a right surangular, twelve isolated dentary teeth, four cervical vertebrae, a distal caudal vertebra, three cervical ribs, and a coracoid. This specimen is stored in the Museum of Nature and Human Activities, Hyogo, Sanda City, Hyogo Prefecture, Japan. Diagnosis: A hadrosaurid with unique characters in having only a single tooth as a minimum number of functional teeth per tooth position in the middle of the dentary dental battery (9th, 11th, 14th, 16th, 19th, 21st, and 23rd tooth positions) and the complete absence of the “branched ridges” on the dentary tooth occlusal surfaces. It is also unique in the combination of the additional following characters: the low angle between the dentary symphysis and lateral surface of the dentary and ventrally facing ventral surface of the surangular. Etymology: “Yamato” refers to the ancient name for a region of the Japanese archipelago, including the western half of the main island (Honshu Island), Shikoku Island, and the northern half of Kyushu Island, ruled by the Yamato Kingdom from the third to the seventh century. “Sauros” means reptiles. The specific name, “izanagi”, refers to a deity in Japanese mythology, which created eight countries of Yamato with another deity, Izanami, based on the oldest history book in Japan called “Kojiki (Records of Ancient Matters)”, published in 712 CE (Common Era). The first country created was the Awaji Island, followed by the Shikoku, Oki, Kyushu, Iki, Tsushima, Sado, and Honshu islands. Locality and horizon: Locality Aw (Morozumi) of Yura Town, Sumoto City of the Awaji Island, Hyogo Prefecture, Japan; the early Maastrichtian (71.94–71.69 Ma) Kita-ama Formation of the Izumi Group.    Yoshitsugu Kobayashi, Ryuji Takasaki, Katsuhiro Kubota and Anthony R. Fiorillo. 2021. A New Basal Hadrosaurid (Dinosauria: Ornithischia) from the latest Cretaceous Kita-ama Formation in Japan implies the Origin of Hadrosaurids. Scientific Reports. 11: 8547. DOI: 10.1038/s41598-021-87719-5 | ||||||||||
| 3:32a | [Ichthyology • 2021] Etelis boweni • A New Cryptic Deepwater Eteline Snapper (Perciformes: Lutjanidae) from the Indo‐Pacific
Abstract A new species of Etelis is described based on 16 specimens collected from the Red Sea and Western Australia, with confirmed genetic records throughout the Indo‐West Pacific. It is similar to and was often misidentified as Etelis carbunculus Cuvier, with both species sharing the diagnostic character of low number of developed gill rakers. Nonetheless, the two species are genetically divergent and differ morphologically in adult body length; proportions of eye, snout, cheek and caudal fin; shape of head, opercular spine and sagittal otolith; and coloration of the tip of the upper caudal fin. Etelis boweni has a wide Indo‐west Pacific distribution that largely overlaps with E. carbunculus, and the two species are often caught on the same fishing line. Keywords: Actinopterygii, cryptic species, ichthyology, Pisces, snappers, taxonomy    Etelis boweni sp. nov. Bowen's Red Snapper, Giant Ruby Snapper Etymology: The authors are grateful to name this fish after Dr. Brian W. Bowen of the Hawaiʻi Institute of Marine Biology, University of Hawaiʻi, in recognition of his contributions to the field of Ichthyology, and in particular for his use of molecular genetic techniques in support of the taxonomy of fishes.
Kimberly R. Andrews, Iria Fernandez‐Silva, John E. Randall and Hsuan‐Ching Ho. 2021. Etelis boweni sp. nov., A New Cryptic Deepwater Eteline Snapper from the Indo‐Pacific (Perciformes: Lutjanidae). Journal of Fish Biology. DOI: 10.1111/jfb.14720 |








| << Previous Day |
2021/04/28 [Calendar] |
Next Day >> |