|
|
| Пишет ivanov_petrov ( @ 2009-04-25 08:19:00 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Революция в зоологии: новая система билатерий
В.В. Малахов опубликовал статью в "Природе" №3
http://elementy.ru/lib/430793
Разговор о новой старой системе... То есть многие таксоны вроде Экдисозой давние уж, и споры давние, И система новая - это объединение данных молекулярной филогенетики. Вместо старых сравнительноанатомических данных.
Старая ср-анат система уходит в никуда. Примерно как система жизненных форм Теофраста исчезли... сменившись системой таксонов, а жизненные формы потом стали делать отдельно и уже не так рьяно, так и тут - видно, что систему выстраивают на молекулярных признаках, то, что было сравнительной анатомией, теперь мыслится как нечто вроде "уровней организации" и прекращает бытие в регулярном качестве. Дело в том, что пока это считалось системой - многие люди регулярно отсматривали всякое зверье и искали противоречия. Как только объявляется сезон вольной охоты и старая система уходит в "данные сравнительной анатомии" - это никто особенно проверять не будет и что там будет скапливаться... понять можно будет лишь с приложением чрезвычайных усилий.
Это мои замечания. Малахов же подчеркивает преемственность - мол, вторичноротые-то остались, и новые объяснения, которых раньше сделать не могли.

(картинка только для иллюстрации и вольных ассоциаций - не из текста Малахова)
Классическая система - от общего предка билатерий отходят "паренхиматозные черви" (турбеллярии, плоские черви, немертины),



потом от общего ствола отходят первичнополостные черви, нематоды, головохоботные, коловратки,



потом возникают все целомические - веером и сразу: членистоногие, трохофорные (кольчатые, моллюски, сипункулиды), щупальцевые (форониды, мшанки, брахиоподы) и вторичноротые (полухордовые, иглокожие, хордовые).
А в новой системе: общий предок билатерий дает вторичноротых (сразу!) и сеcтринская группа - все остальные. Потом от остальных отделяются Линяющие (членистоногие, нематоды, головохоботные). И остается две группы, сестринские трохофорные (плоские черви, кольчатые, моллюски, сипункулиды) и с ними в паре щупальцевые.
Вот такая загогулина.

Обращается внимание на то, что у турбеллярий сложная половая система - не особенно то, мол, примитивны. А у целомических - наружное оплодотворение, самое примитивное... У первичноротых - особенное спиральное дробление. А у вторичноротых - более примитивное недетерминированное (радиальное) дробление. А у членистоногих нет целома. И т.п. - известные странности классической системы, на каждую странность полагалось свое объяснение.
новая же система...
По молекулярным данным щупальцевые близки трохофорным. Новый таксон лофотрохофорных. Предки билатерий - вендские кишечнополотные, ползающие по грунту. (То есть вендскую фауну автор решительно относит к кишечнополостным и никаких). Энтероцельная организация целома из карманов кишечника. Кодекс примитивности: сквозной кишечник, сегментированный целом, метамерные конечности, простая половая система, наружное оплодотворение, радиальное дробление.

Про развитие хордовых - автор напоминает факт http://www.soil.msu.ru/~invert/main

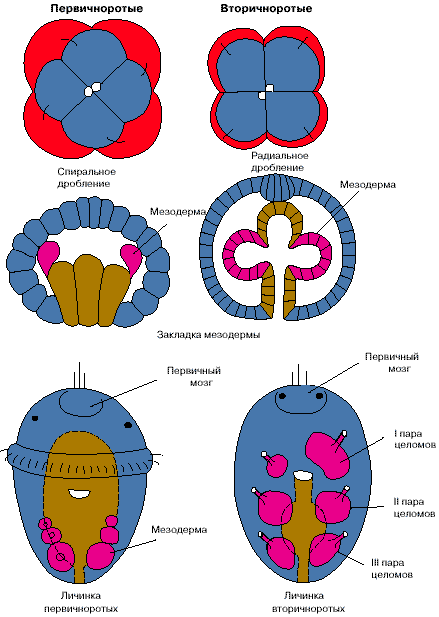
Сравнение планов строения различных групп вторичноротых (личинка асцидий и ланцетник изображены в перевернутом положении). 1 - целомы первой пары, 2 - целомы второй пары, 3 - целомы третьей пары, 4 - кровеносная система, 5 - нервная система. Стрелки показывают направление тока крови.
Теперь Линяющие. Выход на сушу, водорослевые маты и прочая лирика про плащевой сток с континентов без русел и пр. Кутикула с хитином, не растягивается - потому линьки, которые у всех обеспечены гормоном группы экдизонов. Удивительная синапоморфия.

Далее автор говорит о разных планах строения линяющих. И тут (мне кажется) неприятная засада. Ясное дело, если строить филогении по разным системам органов (признаков) - будут противоречия. Всегда. Это не исключение, а правило, решаются они различно, но всегда в общем случае - через придание весов. Мол, это сходство не важно, а то важно. А тут говорится - ну как же могут независимо возникнуть экдизоны в нескольких разных группах, вероятность случайного возникновения чудовищно мала. это потрясающе. что этот гормон маркирует группы единого происхождения... А планы строения могут случайно возникнуть разные? Генетичесое обеспечение плана строения известно и это - очень короткая и легко возникающая de novo последовательность? Здесь - в изложении - допущен сбой в логике. Просто решено признаку наличия экдизона, комплексу линьки - придавать большой вес. А почему? В рамках статьи в Природе ответить и нельзя. Но мне по неграмотности - не ясно.
Далее рассказ о целоме. Целом нужен как опорная система в теле. Но раз есть внешний скелет - нафиг такую систему, и потому у всяких членистоногих целом разрушается. Полость тела Линяющих и в т.ч. членистоногих - это первичная полость, гемоцель. Это гомологично полостям наших кровеносных сосудов. У Линяющих вся полость тела - гемоцель, и есть маленькие участки целома вокруг половых желез и органов выделения, это выворотка наизнанку от вторичноротых, у которых наоборот вся полость тела - целом, и тонкие труборчки кровеносных сосудов - гемоцель, первичная полость. Отличия этих полостей? Целом выстлан клетками, а гемоцель - неклеточной базальной пластинкой. (красота... насекомые - как маленькие частички кровеносных сосудов, самоползающие).

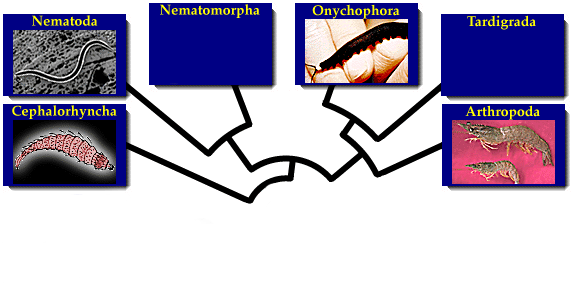
А круглые черви? Они произошли от членистоногих. Те зарылись в землю, потеряли конечности - и пожалте бриться - аскарида. А более глубокие предки были не членистоногие, скорее напоминали онихофор.
Палеосколециды - похожи на головохоботных (приапулиды, лорициферы, киноринхи, волосатики) - промежуточная группа между лобоподами и головохоботными.



Нематоды произошли педоморфозом. Это счиается универсальным механизмом возникновения множества крупных таксонов - с измельчанием и сменой плана строения. Вторично могоут укрупняться. Так и круглые черви - от типа многоножки - личинки, без ног - вот и немматоды.
У Линяющих нет ресничных личинок. Науплиусы ракообразных и личинки насекомых - это во=торичные личинки, передвигаются с помощью конечностей. Эмбриогенез Линяющих так изменен, что не поймешь, откуда они. Среди членистоногих ракообразные и насекомых поделили сушу и море. Раки не пустили насекомых в море - заняв там все экологические ниши, подходящие членистоногим. (Хм.Хм. Ничо так объяснение, решительное)

Щупальцевые Lophophorata - примитивные лофотрохофорные Lophotrochozoa. Предки их были сегментированы, метамерные придатки-конечности. Параподии. Важнейший их признак - щетинки на параподиях. У всех трохофорных строение щетинок одинаково. Стержень щетинки формируется всего одной клеткой. И опять - вот так объединяют группу, а потом говорят - в этом стволе разошлись разные планы строения..... Опять взвешивание, и в статье не сказано - по каким параметра. причем ясно. что тех прищзнаков много. и дают они лишь частично разрешенные деревья. И все это, говоря проще, вилами на воде - а туда же... Впрочем, увлекся. Прошу простить.
Далее о щупальцевых - Brachiopoda, Phoronida, Bryozoa. Это целая поэма, как они свой план строения вырабатывали. У плеченогих раковина из двух створок. Как у моллюсков. Но у моллюсков створки слева и справа от тела. А у плеченогих - это две створки, обе спинные, передняя и задняя спинная. тварь - предполагается - согнулась пополам, как бы втянув живот, и образовалась створка на спинной стороне ближе к голове и дальше от головы. Щупальцевые - фильтраторы, эту группу выделяют по лофофору, фильтрационному аппарату.




Трохофорные - со спиральным дроблением в эмбриогенезе. Тут опираются на план строения личинки. У многих перистальтическая локомоция. Мелкие трохофорные - группы архианнелид, коловратки, гастротрихи, микрогнатозои - все все они - педогенезом произошли от личинок вымерших крупных трохофорных. Если совсем неправильно - скажем так, от личинок моллюсков.. (этого Малахов не говорит, я напоминаю, кто такие трохофорные).



Тупбеллярии произошли тоже педоморфозом.
В общем, мне кажется, что в этой системе педоморфоз играет роль универсальной отмычки. Есть масса вопросов при создании системы, и многие из них решаются - попросту указанием на педоморфоз, от которого и следов-то почти не остается. Не сказал бы, что это очень изящное решение.
В завершение статьи говорится о нерешенных проблемах новой системы. Что с первичным ртом? Щупальцевые отнесены к первичноротым - и теперь оба ствола билатерий - и первично-, и вторичноротые - могут иметь радиальное дробление и энтероцельную закладку целома. Нет места морским стрелкам, ксенотурбеллидам. Не выделены бескишечные турбеллярии.
(Я бы в общем сказал так: была система, наследующая системе В.Н. Беклемишева - основанная на планах строения и симметричных преобразованиях. Потом долго копали, уточняли и разрушали деталями, докопались - черты планов строения принесены в жертву частным признакам, от этого старая система разрушена. Я бы сказал, что новой - не появилось. Хлам это, а не система - хоть это и пустые слова, поскольку для филогенетика никакой системной организованости и не предполагается, любой набор считается осмысленным).
Важно понимать - это сделано не по целым геномам. Выделили небольшое число групп генов, которые пока считаются важными, построили деревья для этих немногих групп. Полученное расшифровали примерно вот так. Когда данные будут уточнены - может измениться решительно всё.
Так что пока так - для сторонних людей

это не система Малахова. той в электронном виде нет, это близкая
в качестве шутки - вот представляют эволюцию будущего...

В.В. Малахов опубликовал статью в "Природе" №3
http://elementy.ru/lib/430793
Разговор о новой старой системе... То есть многие таксоны вроде Экдисозой давние уж, и споры давние, И система новая - это объединение данных молекулярной филогенетики. Вместо старых сравнительноанатомических данных.
Старая ср-анат система уходит в никуда. Примерно как система жизненных форм Теофраста исчезли... сменившись системой таксонов, а жизненные формы потом стали делать отдельно и уже не так рьяно, так и тут - видно, что систему выстраивают на молекулярных признаках, то, что было сравнительной анатомией, теперь мыслится как нечто вроде "уровней организации" и прекращает бытие в регулярном качестве. Дело в том, что пока это считалось системой - многие люди регулярно отсматривали всякое зверье и искали противоречия. Как только объявляется сезон вольной охоты и старая система уходит в "данные сравнительной анатомии" - это никто особенно проверять не будет и что там будет скапливаться... понять можно будет лишь с приложением чрезвычайных усилий.
Это мои замечания. Малахов же подчеркивает преемственность - мол, вторичноротые-то остались, и новые объяснения, которых раньше сделать не могли.
(картинка только для иллюстрации и вольных ассоциаций - не из текста Малахова)
Классическая система - от общего предка билатерий отходят "паренхиматозные черви" (турбеллярии, плоские черви, немертины),
потом от общего ствола отходят первичнополостные черви, нематоды, головохоботные, коловратки,
потом возникают все целомические - веером и сразу: членистоногие, трохофорные (кольчатые, моллюски, сипункулиды), щупальцевые (форониды, мшанки, брахиоподы) и вторичноротые (полухордовые, иглокожие, хордовые).
А в новой системе: общий предок билатерий дает вторичноротых (сразу!) и сеcтринская группа - все остальные. Потом от остальных отделяются Линяющие (членистоногие, нематоды, головохоботные). И остается две группы, сестринские трохофорные (плоские черви, кольчатые, моллюски, сипункулиды) и с ними в паре щупальцевые.
Вот такая загогулина.
Обращается внимание на то, что у турбеллярий сложная половая система - не особенно то, мол, примитивны. А у целомических - наружное оплодотворение, самое примитивное... У первичноротых - особенное спиральное дробление. А у вторичноротых - более примитивное недетерминированное (радиальное) дробление. А у членистоногих нет целома. И т.п. - известные странности классической системы, на каждую странность полагалось свое объяснение.
новая же система...
По молекулярным данным щупальцевые близки трохофорным. Новый таксон лофотрохофорных. Предки билатерий - вендские кишечнополотные, ползающие по грунту. (То есть вендскую фауну автор решительно относит к кишечнополостным и никаких). Энтероцельная организация целома из карманов кишечника. Кодекс примитивности: сквозной кишечник, сегментированный целом, метамерные конечности, простая половая система, наружное оплодотворение, радиальное дробление.
Про развитие хордовых - автор напоминает факт http://www.soil.msu.ru/~invert/main
Сравнение планов строения различных групп вторичноротых (личинка асцидий и ланцетник изображены в перевернутом положении). 1 - целомы первой пары, 2 - целомы второй пары, 3 - целомы третьей пары, 4 - кровеносная система, 5 - нервная система. Стрелки показывают направление тока крови.
Теперь Линяющие. Выход на сушу, водорослевые маты и прочая лирика про плащевой сток с континентов без русел и пр. Кутикула с хитином, не растягивается - потому линьки, которые у всех обеспечены гормоном группы экдизонов. Удивительная синапоморфия.
Далее автор говорит о разных планах строения линяющих. И тут (мне кажется) неприятная засада. Ясное дело, если строить филогении по разным системам органов (признаков) - будут противоречия. Всегда. Это не исключение, а правило, решаются они различно, но всегда в общем случае - через придание весов. Мол, это сходство не важно, а то важно. А тут говорится - ну как же могут независимо возникнуть экдизоны в нескольких разных группах, вероятность случайного возникновения чудовищно мала. это потрясающе. что этот гормон маркирует группы единого происхождения... А планы строения могут случайно возникнуть разные? Генетичесое обеспечение плана строения известно и это - очень короткая и легко возникающая de novo последовательность? Здесь - в изложении - допущен сбой в логике. Просто решено признаку наличия экдизона, комплексу линьки - придавать большой вес. А почему? В рамках статьи в Природе ответить и нельзя. Но мне по неграмотности - не ясно.
Далее рассказ о целоме. Целом нужен как опорная система в теле. Но раз есть внешний скелет - нафиг такую систему, и потому у всяких членистоногих целом разрушается. Полость тела Линяющих и в т.ч. членистоногих - это первичная полость, гемоцель. Это гомологично полостям наших кровеносных сосудов. У Линяющих вся полость тела - гемоцель, и есть маленькие участки целома вокруг половых желез и органов выделения, это выворотка наизнанку от вторичноротых, у которых наоборот вся полость тела - целом, и тонкие труборчки кровеносных сосудов - гемоцель, первичная полость. Отличия этих полостей? Целом выстлан клетками, а гемоцель - неклеточной базальной пластинкой. (красота... насекомые - как маленькие частички кровеносных сосудов, самоползающие).
А круглые черви? Они произошли от членистоногих. Те зарылись в землю, потеряли конечности - и пожалте бриться - аскарида. А более глубокие предки были не членистоногие, скорее напоминали онихофор.
Палеосколециды - похожи на головохоботных (приапулиды, лорициферы, киноринхи, волосатики) - промежуточная группа между лобоподами и головохоботными.
Нематоды произошли педоморфозом. Это счиается универсальным механизмом возникновения множества крупных таксонов - с измельчанием и сменой плана строения. Вторично могоут укрупняться. Так и круглые черви - от типа многоножки - личинки, без ног - вот и немматоды.
У Линяющих нет ресничных личинок. Науплиусы ракообразных и личинки насекомых - это во=торичные личинки, передвигаются с помощью конечностей. Эмбриогенез Линяющих так изменен, что не поймешь, откуда они. Среди членистоногих ракообразные и насекомых поделили сушу и море. Раки не пустили насекомых в море - заняв там все экологические ниши, подходящие членистоногим. (Хм.Хм. Ничо так объяснение, решительное)
Щупальцевые Lophophorata - примитивные лофотрохофорные Lophotrochozoa. Предки их были сегментированы, метамерные придатки-конечности. Параподии. Важнейший их признак - щетинки на параподиях. У всех трохофорных строение щетинок одинаково. Стержень щетинки формируется всего одной клеткой. И опять - вот так объединяют группу, а потом говорят - в этом стволе разошлись разные планы строения..... Опять взвешивание, и в статье не сказано - по каким параметра. причем ясно. что тех прищзнаков много. и дают они лишь частично разрешенные деревья. И все это, говоря проще, вилами на воде - а туда же... Впрочем, увлекся. Прошу простить.
Далее о щупальцевых - Brachiopoda, Phoronida, Bryozoa. Это целая поэма, как они свой план строения вырабатывали. У плеченогих раковина из двух створок. Как у моллюсков. Но у моллюсков створки слева и справа от тела. А у плеченогих - это две створки, обе спинные, передняя и задняя спинная. тварь - предполагается - согнулась пополам, как бы втянув живот, и образовалась створка на спинной стороне ближе к голове и дальше от головы. Щупальцевые - фильтраторы, эту группу выделяют по лофофору, фильтрационному аппарату.
Трохофорные - со спиральным дроблением в эмбриогенезе. Тут опираются на план строения личинки. У многих перистальтическая локомоция. Мелкие трохофорные - группы архианнелид, коловратки, гастротрихи, микрогнатозои - все все они - педогенезом произошли от личинок вымерших крупных трохофорных. Если совсем неправильно - скажем так, от личинок моллюсков.. (этого Малахов не говорит, я напоминаю, кто такие трохофорные).
Тупбеллярии произошли тоже педоморфозом.
В общем, мне кажется, что в этой системе педоморфоз играет роль универсальной отмычки. Есть масса вопросов при создании системы, и многие из них решаются - попросту указанием на педоморфоз, от которого и следов-то почти не остается. Не сказал бы, что это очень изящное решение.
В завершение статьи говорится о нерешенных проблемах новой системы. Что с первичным ртом? Щупальцевые отнесены к первичноротым - и теперь оба ствола билатерий - и первично-, и вторичноротые - могут иметь радиальное дробление и энтероцельную закладку целома. Нет места морским стрелкам, ксенотурбеллидам. Не выделены бескишечные турбеллярии.
(Я бы в общем сказал так: была система, наследующая системе В.Н. Беклемишева - основанная на планах строения и симметричных преобразованиях. Потом долго копали, уточняли и разрушали деталями, докопались - черты планов строения принесены в жертву частным признакам, от этого старая система разрушена. Я бы сказал, что новой - не появилось. Хлам это, а не система - хоть это и пустые слова, поскольку для филогенетика никакой системной организованости и не предполагается, любой набор считается осмысленным).
Важно понимать - это сделано не по целым геномам. Выделили небольшое число групп генов, которые пока считаются важными, построили деревья для этих немногих групп. Полученное расшифровали примерно вот так. Когда данные будут уточнены - может измениться решительно всё.
Так что пока так - для сторонних людей
это не система Малахова. той в электронном виде нет, это близкая
в качестве шутки - вот представляют эволюцию будущего...