[Recent Entries][Archive][Friends][User Info]
December 4th, 2011
| December 4th, 2011 | |
|---|---|
| 01:12 pm [industrialterro] [Link] |
Styracocephalus Стиракоцефал (Styracocephalus platyrhynchus) – примитивный терапсид из средней перми (средний горизонт зоны Tapinocephalus) Южной Африки. Обычно считается тапиноцефалом, но некоторыми исследователями относится к бурнетиидам. Животное крупных размеров (длиной, вероятно, до 2,5 метров). Морда очень длинная, низкая. Мощные выросты над глазами, развитые «рога» табулярных костей, направленные назад, боковые выросты на чешуйчатых костях. Зубной край верхней челюсти прямой, нижняя челюсть массивная с высоким симфизом. Развиты мощные резцы, сохраняются увеличенные клыки, заклыковые зубы многочисленные (до 11 пар). Небные зубы слабо развиты. Животное растительноядное. Описан С. Хоутоном в 1929 году. Долгое время был известен лишь по одному разрушенному черепу, но в 1997 году было опубликовано дополнительное описание еще нескольких черепов. В отличие от бурнетиид и эстемменозухов, выросты черепа образованы другими костями. Поэтому, вероятно, представляет особую ветвь дейноцефалов. С другой стороны, недавно стиракоцефала вновь сблизили с эстемменозухами и бурнетиаморфами.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 01:47 pm [industrialterro] [Link] |
Suminia Суминия (Suminia getmanovi) — доисторический синапсид, относившийся к кладу Anomodontia, живший около 260 млн лет назад в позднем пермском периоде («зона II»). Известный экземпляр был обнаружен в г. Котельнич на реке Вятка в песчаниковых отложениях: последнее говорит о том, что скорее всего суминия была жителем речной дельты. Возможно, животное обитало и в других зонах, однако других окаменевших останков не сохранилось. Зубы обнаруженного экземпляра были сильно изношены, что говорит о том, что она питалась растительной пищей с большим содержанием частиц песка. Образцы, найденные позднее, говорят о том, что суминия вела древесный образ жизни. Единого взгляда на систематическое положение суминии нет. Наталия Рыбчински в работе, посвященной анатомии черепа и филогенетической позиции Suminia getmanovi, помещает род Suminia в инфраотряд Venyukovioidea Watson & Romer, 1956. М. Ф. Ивахненко рассматривает суминию в рамках семейства Galeopidae Broom, 1912 (инфраотряд Dromasauria, подотряд Dicynodontia). Вид Suminia getmanovi найден в местонахождении Котельнич, Котельничский р-н, Кировской обл. (верхняя пермь, татарский отдел, верхнесеверодвинский подъярус, северодвинский горизонт). Входит в состав котельничской фауны (котельничский фаунистический комплекс). Типовой экземпляр — полный скелет хорошей сохранности. В дальнейшем было найдено множество экземпляров, в том числе крупное скопление скелетов (захоронение из останков 15 животных). Ископаемые остатки нередки, представлены несколькими скелетами и черепами, а также изолированными зубами. Остатки суминий неустановленной видовой принадлежности и определенные как S. cf. getmanovi известны и из других местонахождений. Зубные кости, определенно принадлежащие роду Suminia, известны из местонахождения Потеряха-2, Нюксенский р-н, Вологодская обл. (северодвинский горизонт, верхнесеверодвинский подъярус, татарский отдел, верхняя пермь). Предчелюстная кость, отличающаяся деталями строения зубов от типового вида, найдена в местонахождении Устье Стрельны, Великоустюгский р-н, Вологодская обл. (северодвинский горизонт, верхнесеверодвинский подъярус, татарский отдел, верхняя пермь). Совершенно неожиданный взгляд на образ жизни суминии высказан в одной из недавних работ. Йорг Фребиш (Jörg Fröbisch) из Музея Филда в Чикаго (Field Museum in Chicago) и Роберт Рейц (Robert Reisz) из Университета Торонто в Канаде (University of Toronto), исследовав окаменелые скелеты суминии, обнаружили у этого аномодонта несколько адаптаций к древесному образу жизни. Рейц и Фребиш установли, что суминия была, возможно, древесным животным, и что она является на сегодняшнее время самым ранним известным древесным позвоночным. Она поднялась на деревья за 30 миллионов лет до похожих на хамелеонов дрепанозаврид, занимавших аналогичную нишу. Исследованный палеонтологами материал состоял из одного блока, содержащего остатки скелетов более десяти животных. Хотя такие захоронения могут быть трудными для изучения, они также чрезвычайно информативны, особенно с учетом возможности изучения различий между разными животными одного вида из одного места. Скелеты на плите принадлежали животным разного возраста, от полувзрослых до взрослых, а сохранность костей свидетельствует о быстром захоронении, вызванным какой-то небольшой катастрофой. Древесные животные редки в ископаемой летописи, поскольку они не живут в местах с благоприятными условиями для образования окаменелостей, поэтому ископаемые остатки суминии являются неожиданностью для палеонтологов. Прекрасная сохранность материала позволила Рейцу и Фребишу изучить анатомию суминии во всех деталях. По мнению ученых, наиболее любопытной особенностью являются кисти и стопы животных. Кисть и стопа большие по сравнению с остальной конечностью. Кисть суминии сильно отличается от кисти наземных аномодонтов и больше похожа на кисти других древесных позвоночных из разных групп и периодов времени. Большая часть длины кисти и стопы образована длинными тонкими пальцами с удлиненными предпоследними фалангами, которые по пропорциям и форме сходны с таковыми у некоторых современных животных, включая шерстокрылов, крыланов и ящериц, цепляющихся когтями при лазании по деревьям. Первые пальцы кисти и стопы могли отгибаться под углом к остальным четырем пальцам, и, возможно, использовались как противопоставленные хватательные пальцы («большие пальцы»). Это механизм, появление которого можно было бы ожидать у животного, обхватывающего конечностями ветви деревьев при перемещении в кроне и, по-видимому, кисти и пальцы суминии в этом отношении аналогичны кистям и пальцам некоторых древесных животных. Более мелкие особенности также указывают на древесный образ жизни. Хвост суминии относительно длинный, а хвостовые позвонки первой половины хвоста несут сильные отростки. Эти отростки, возможно, поддерживали мускулы, которые позволяли суминии использовать хвост для равновесия или как цепкий хватательный орган. Оказалось, что линии, которые стали вести древесный образ жизни развивали аналогичные приспособления неоднократно, и это сходство объясняется конвергентной эволюцией. Сравнение суминии с другими древесными позвоночными животными подтверждает эту гипотезу, таким образом, Рейц и Фребиш нашли доказательства того, что суминия могла перемещаться по деревьям, цепляясь и обхватывая ветви. Почему предки суминии стали жить на деревьях, однако, пока остается неясным. Суминия, жившая 260 млн. лет назад, является первым известным позвоночным животным с такой степенью специализации к древесному образу жизни. Рейц и Фребиш отмечают, что поздняя пермь и местонахождение Котельнич, где была найдена суминия, предоставляют свидетельства наиболее раннего этапа становления современных наземных экосистем с большим количеством потребителей растений, поддерживающим малое количество главных хищников. В конце пермского периода окружающая среда обеспечивала существование множества растительноядных животных (около 83% всех наземных позвоночных) и гораздо меньшего числа плотоядных (около 13% всех наземных позвоночных). Хотя древесный образ жизни помогал суминиям спасаться от хищников, авторы статьи предполагают, что именно конкуренция с другими растительноядными заставила некоторых синапсид подняться на деревья. Эти растения были ресурсом, не использовавшимся другими позвоночными, что, возможно, и предопределило эволюцию суминии от наземных предков. Таким образом, суминия нашла новый способ использовать листву верхушек деревьев и сделала первый известный шаг в нишу, к которой возвратятся позвоночные животные еще несколько раз за следующие 260 миллионов лет. В местонахождении Котельнич были найдены небольшие копролиты овально-яйцевидной формы, длиной 1-2 см, с углистым материалом внутри. В копролитах обнаружены фрагменты кутикулы, возможно, принадлежавшей птеридоспермам (семенным папоротникам), а также фрагменты древесины. Эти копролиты могли принадлежать Suminia getmanovi. Блок со скелетами суминий, изученный Фребишем и Рейцем, также содержал мелкие (менее 10 мм) копролиты, внутри которых обнаружены фрагменты листьев.
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 02:02 pm [industrialterro] [Link] |
Sycosaurus Sycosaurus is an extinct genus of Gorgonopsia. It was first named by Haughton in 1924, and contains four species, S. brodiei, S. kingoriensis, S. laticeps, S. vanderhorsti. Sycosaurus's skull is about medium size for a Gorgonopsid. It has a dental formula of i5, c1, pc5.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 02:13 pm [industrialterro] [Link] |
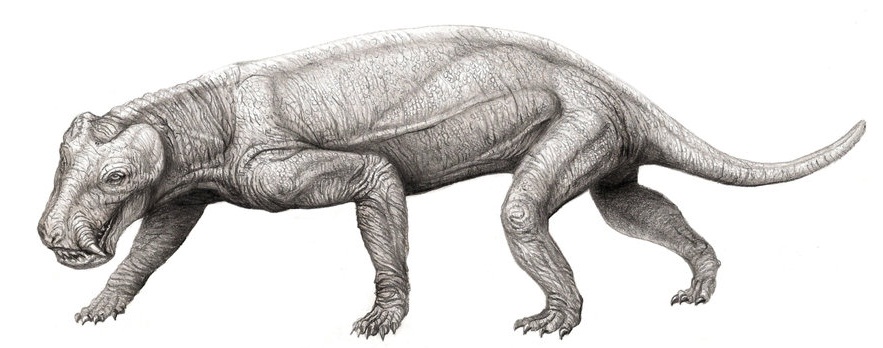
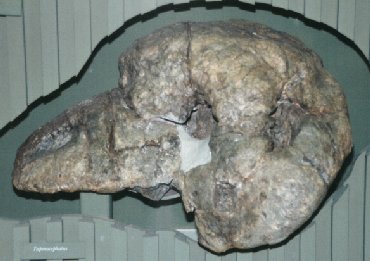
Tapinocephalus Tapinocephalus is a genus of large herbivorous dinocephalian that lived during the Middle Permian Period. These stocky, barrel-bodied animals were characterised by a massive bony skull roof and short weak snout. It is thought that, like the rest of the members of its family, the animals engaged in head-butting intraspecific behavior, possibly for territory or mates. The fossil remains (skull and postcranial elements) of Tapinocephalus are known from the Lower, Middle, and Upper part of the Tapinocephalus Assemblage Zone ( Capitanian age) of the Lower Beaufort Beds of the South African Karoo. Only the type species, T. atherstonei is now considered valid for this genus. In life, these animals were over 3 meters (10 ft) in length and weighed around 1.5 to 2 metric tonnes (1.6 to 2 short tons), making them among the largest animals of their time. Keratocephalus and Moschops are closely related contemporary forms, of similar size and habits. Тапиноцефалы (Tapinocephalia) — подотряд растительноядных дейноцефалов. Известны из средней перми Южной Африки, России и Китая. Включают два надсемейства — титанозухи и собственно тапиноцефалы. Сюда же может относиться проблематичный род стиракоцефал (Styracocephalus) из средней перми Южной Африки, которого иногда объединяют с бурнетиидами. Собственно тапиноцефалы (надсемейство Tapinocephaloidea) включает единственное семейство Tapinocephalidae. Это очень крупные животные (весом до 1 −1,5 тонн). Тело массивное, округлое, хвост очень короткий. Конечности очень массивные, передние чуть длиннее задних. Вероятно, при жизни передняя половина тела была приподнята (возможно, это указывает на питание какой-то приподнятой над почвой растительностью). Морда при обычном положении головы опущена несколько вниз (затылок резко скошен назад). Череп очень массивный, с чрезвычайно развитым пахиостозом в надглазничной и лобно-теменной области. Толщина костей черепа у некоторых видов достигала 30 см. Глазницы и височные впадины небольшие, височные впадины открыты сверху. Зубы немногочисленные, резцы обычно очень мощные. Небных зубов нет. Клыки выражены лишь у самых примитивных форм. Предполагается, что пахиостоз костей черепа был необходим для внутривидовых схваток (как у баранов или горных козлов). Тем не менее, это оспаривается отечественными учеными, которые указывают, что кости лба не были покрыты роговыми образованиями и служили для терморегуляции (их пронизывали кровеносные сосуды). Шкура тапиноцефалов, вероятно, была мягкой и лишенной чешуй, с многочисленными железами. На лицевой части черепа у некоторых видов обнаружены желобчатые углубления, возможно, вмещавшие кожные железы. Известны отпечатки следов из Южной Африки. Тапиноцефалы были растительноядными, причем для измельчения пищи они использовали мощные резцы, снабженные пяткообразными выступами. Основу их питания могли составлять мертвые стволы каламитов, которые накапливались по берегам водоемов. Самый примитивный тапиноцефал — Tapinocaninus pamelae из зоны Eodicynodon Южной Африки. Он сохранял заметные верхние клыки, но в остальном очень сходен с более поздними родичами. Найденный в Приуралье улемозавр представляет родственную примитивную форму. Недавно показано, что приуральские дейтерозавры также являются примитивными тапиноцефалами. Более прогрессивные тапиноцефалы включают примерно 12 — 13 родов преимущественно из Южной Африки. Истинное количество родов неизвестно, так как многие из них оказались синонимами, поскольку описывались на основании остатков разнополых и разновозрастных особей одного и того же вида. Tapinocephalinae — короткомордые тапиноцефалы с относительно слабыми зубами. Пахиостоз чрезвычайно развит, иногда в виде куполообразных выростов на крыше черепа. Наиболее известны мосхопс (Moschops), тапиноцефал (Tapinocephalus), кератоцефал (Keratocephalus). Самый крупный вид — Tapinocephalus atherstonei, длиной до 4 метров. Мосхопса и его ближайших родичей (Criocephalus, Avenantia, Delphinognathus) иногда выделяют в отдельное подсемейство.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 02:29 pm [industrialterro] [Link] |
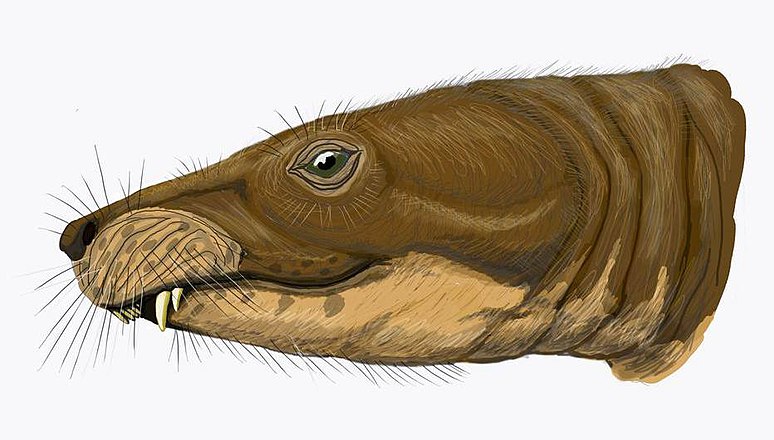
Theriognathus Териогнат (Theriognathus) – позднепермский тероцефал из семейства вайтсиид. Известен также под названиями Aneugomphius, Whaitsia, Notosollasia и др. В реальности все эти тероцефалы оказались представителями не только одного рода, но, вероятно, и одного вида - Theriognathus ictidoceps (или T. microps). Длина черепа от 8 до 31 см, общая длина до 2 метров. Морда относительно широкая, череп низкий. Хорошо развиты резцы, присутствуют 2 пары верхних клыков (вторая пара может быть замещающей). Верхние резцы нависают над нижними, направлены вниз и назад. Заклыковые зубы обычно отсутствуют. Твердого вторичного неба нет, но мягкое небо могло продолжаться далеко назад, помогая дышать во время еды. Предполагается, что вместо задних зубов могли быть роговые пластины, покрывавшие десны. Скелет массивный. Считается всеядным животным, но известна находка черепа (отнесенного к роду Notosollasia) с застрявшим между челюстей поврежденным черепом мелкого дицинодонта. Это позволяет предположить, что териогнаты были хищниками, которые при атаке хватали жертву за голову (так делает в наше время ягуар). Возможно, мелкая добыча проглатывалась целиком. Известен из позднепермских отложений (зоны Cistecephalus - Dicynodon) Южной Африки.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 03:09 pm [industrialterro] [Link] |
Titanophoneus Титанофонеус (Titanophoneus) — хищный дейноцефал группы антеозавров. Известен из «средней» перми Приуралья и Поволжья (описан из Ишеево в Татарии — нижнетатарский ярус). Во многом сходен с южноафриканским антеозавром. Череп высокий и узкий, у взрослых особей развиты утолщения заглазничной и лобной области (пахиостоз). Мощные резцы и клыки, 8—9 пар мелких заклыковых зубов. Ноги мощные, не очень длинные, с широкими кистями и стопами (возможно, имелась плавательная перепонка). Хвост длинный. В целом скелет довольно легкий. Молодые особи (первые из описанных экземпляров) достигали в длину 3 метров, черепа взрослых могли достигать 60 см в длину. Типовой вид — T. potens, описан И. А. Ефремовым в 1938 году. Типовой череп (молодая особь) сломан и его передняя часть резко вздернута вверх (именно так он изображался в старых книгах). В реальности он был не столь «бульдогообразным». К данному роду и виду может принадлежать и другой хищный дейноцефал из Ишеево — долиозавриск. М. Ф. Ивахненко обосновывает синонимичность типового вида титанофонеуса и долиозавриска (на чем изначально настаивал И. А. Ефремов). Ископаемые остатки титанофонеев обнаружены в ряде местонахождений Приуралья и Поволжья и датируются серединой поздней перми (уржумский горизонт, нижнетатарский подъярус, уржумский ярус, биармийский отдел). Остатки встречаются довольно часто — к примеру, в коллекции Ишеево они составляют до 15% всех находок, в том числе отсюда известно 2 более или менее полных скелета. Титанофонеи являются одними из лучше всего изученных представителей группы. Они представляли собой крупных дейноцефалов с большой головой, короткой мощной шеей, удлиненным приземистым туловищем, расставленными в стороны сильными конечностями и длинным хвостом. Черепа взрослых животных могли достигать 60 см в длину при общей длине тела около 5 м. В своих фаунистических сообществах они были наиболее крупными хищниками. Череп умеренно высокий, суженный спереди и расширенный в скуловой области, треугольная в сечении лицевая часть несколько удлинена (посторбитальное расстояние укладывается в общую длину черепа 4,5-5 раз). Край верхней челюсти впереди клыков полого поднимается вверх, не образуя резкого уступа. Подобная «бульдогообразность» присутствовала уже у раннепермских пеликозавров и, очевидно, связана со способом хватания добычи. Ноздри и орбиты несколько приподнятые. Глазницы сравнительно небольшие, округлые; глазные впадины направлены косо в стороны и довольно сильно вперед, что было важно при высматривании добычи. Глазное яблоко снабжено кольцом склеротики. Глазничный край скуловой кости с возрастом утолщался. Предлобная кость и верхний край глазницы с бугристой поверхностью. Межглазничный и теменной отделы сужены. Височная дуга высокая, утолщающаяся с возрастом. Задневисочные окна крупные, ямы большие, широко открытые сверху; задний край глубоко вклинен в отросток скуловой кости. Имеется редуцированная надвисочная кость, у старых животных сливающаяся с соседними костями черепа. Межтеменная кость очень высокая, сильно расширенная внизу, с мощным сагиттальным гребнем. Строение задней части черепа очень характерно для антеозавроидей, с отверстием теменного глаза, лежащем на заднем краю на высоком бугре, образованном одними теменными костями. Теменное отверстие крупное, слабо удлиненно-овальное, окруженное высоким костным воротничком; нерв теменного глаза был довольно длинным. Столбик среднего уха (стремечко) имеет вид небольшой распорки в основании черепа между квадратной костью и мозговой коробкой, где он упирается в овальное окно. Это положение столбика как будто бы исключает его подвижность и приспособленность для передачи звуковых колебаний через овальное окно во внутреннее ухо, хотя по своему местоположению он и равнозначен стремечку среднего уха млекопитающих, птиц и пресмыкающихся, имеющих барабанную перепонку. Как воспринимали звук титанофонеи — до конца неясно, но вряд ли животные были глухими. На задней половине носовых костей, предлобных, лобных и заднелобных костях с возрастом развиваются мощные пахиостозные образования с грубой ругозной поверхностью. Пахиостозный бугор назофронтальной области невысокий, захватывает значительную часть носовой кости, полого поднимаясь почти от премаксиллы. Ругозистые утолщения на заглазничной кости высокие, гребневидные. Вся область покрыта сетью борозд кровеносных сосудов. Наличие структур такого типа довольно обычно для растительноядов (например, относительно близких тапиноцефалоидей), но вызывает удивление у хищников — особенно в данном случае, учитывая вытянутую форму черепа. Частично их можно было бы толковать как итог приспособления к большой механической нагрузке со стороны переднего конца морды при хватании и удержании крупной добычи. Однако, у южноафриканских дейноцефалов подобные утолщения намного сильнее и, главное, слишком велики, чтобы в них можно было видеть отражение механической нагрузки. Не объясняет данная версия и странного бугристого строения внешней поверхности костей верхней стороны черепа. Зубная формула: I5/4, С1/1, РС6-8/6-9. Предклыковые зубы крупные, конические, с язычными гребнями и слабыми пятками, в которые упирались вершины зубов противоположной челюсти. В ряде случаев резцы скошены вперед, особенно верхние, очень плотно входящие в промежутки между нижними (как и нижние между верхними). Длинные клыки овальные в сечении, изогнуты назад и обладают мощными, глубоко сидящими корнями. Задний край клыка снабжен мелко зазубренным режущим гребнем, отходящим от вершины зуба к его основанию. На небе имеются особые углубления для вершин нижних клыков. В целом резцы и клыки очень мощные, и, очевидно, хорошо приспособленные к разрыванию даже крупной жертвы. Предклыковые зубы и клыки являлись в первую очередь хватательными, хотя гребни на краях верхних резцов, заходящие за края нижних, делали резцы отчасти и режущими. Наличие на предклыковых зубах зачаточных пяток указывает на их слабо выраженную измельчающую функцию (гораздо более совершенными в этом плане были резцы тапиноцефалоидей). Заклыковые зубы небольшие, слабо согнутые, наклоненные назад, с острыми и уплощенными с боков коронками, обладающими зазубренными режущими кантами. Редкие и сравнительно слабые, режущие по своему типу, они мало годились для измельчения пищи и, скорее всего, играли вспомогательную роль при удержании добычи. Поскольку жевать свою добычу титанофонеи не могли, они глотали ее целиком или отрывали куски мощным рывком головы. Ребра двухголовчатые. Лопатка, ключица и межключица узкие. Лопатка изогнута очень слабо; поскольку ее кривизна при близком расположении к грудной клетке в известной форме отражает ее форму, можно сделать вывод о сжатом с боков туловище (у крокодилов оно обычно уплощенное сверху вниз). Вентральная часть тазового пояса расширенная. В симфизе таза между лобковыми и седалищными костями имеется зона неполного окостенения. В вертлужной впадине между подвздошной, лобковой и седалищной костями окостенение неполное. Подвздошная кость по своему строению промежуточная между таковой древнейших рептилий, где она вытянута назад, и подвздошной костью более прогрессивных синапсид, у которых ее верхний отдел вытянут вперед, как у млекопитающих. Ранее с титанофонеями сближали и описанного по фрагментам черепа адметофонея (Admethophoneus karagalensis). На сегодняшний день его относят к примитивным горгонопсиям типа камагоргона (Kamagorgon ulanovi).
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 03:43 pm [industrialterro] [Link] |
Titanosuchus Titanosuchus ferox ("Fierce Titan crocodile") was a dinocephalian therapsid that lived in the Mid Permian epoch in South Africa. Despite the name, it was not related to crocodiles. Along with its close relatives, Jonkeria and Moschops, Titanosuchus inhabited present-day South Africa around 255 million years ago, in the Late Permian. Titanosuchus was a carnivore and might have eaten both Jonkeria and Moschops, among other vertebrates. Its teeth included sharp incisors and fang-like canines, perfect for biting prey. Titanosuchus rivals that to Titanophoneus, which is also a carnivore and a dinocephalian, but it lived only in Russia. Titanosuchus should not be confused with the therapsid Eotitanosuchus, which belonged to a different family.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 03:58 pm [industrialterro] [Link] |
Ulemica Улемика (Ulemica) — примитивный аномодонт среднепермской эпохи. Близка к венюковиям. Первоначально описана И. А. Ефремовым в 1940 году по черепу из Ишеевского местонахождения как второй вид венюковии (Venyukovia invisa). Долгое время череп изображали в литературе как череп венюковии. Исследования М. Ф. Ивахненко позволили в 1996 году выделить это животное в особый род и в отдельное семейство. Более полный череп из Оренбургской области, изображённый, в частности, в работе П. К. Чудинова как череп типового вида венюковии, принадлежит особому виду улемики — U. efremovi. Длина черепа улемики около 15 см, скелет неизвестен. Остатки встречаются гораздо чаще, чем остатки венюковий — до 15 % находок в Ишееве. Череп по очертаниям напоминает череп дицинодонтов. Нижняя челюсть очень массивная, с лопастевидными выростами на подбородке. Передние резцы мощные, как у грызунов, щёчные зубы сильно расширены. Очень широкая и мощная поверхность предчелюстных, челюстных и зубных костей предполагает ороговение. Посередине верхней челюсти есть один увеличенный зуб, входящий в ямку на нижней челюсти. Боковые движения челюстей при таком строении невозможны, челюсти могли лишь раздавливать пищу. Ефремов считал, что «венюковия» (улемика) питалась подобно грызунам, добывая пищу из твёрдых оболочек. На самом деле, улемики, вероятно, питались твёрдой пищей, которую раздавливали зубами. Пищей могли быть стволики молодых каламитов.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 04:05 pm [industrialterro] [Link] |
Ulemosaurus Улемозавр (Ulemosaurus) — растительноядный дейноцефал из подотряда тапиноцефалов. Обитал в среднепермскую (около 265 млн л.н.) эпоху в Приуралье. Описан А. Н. Рябининым в 1938 году из района села Ишеево в Татарии. В 1980-х годах считался видом африканского рода мосхопс. Тем не менее, улемозавр — более примитивный тапиноцефал, иногда выделяемый в особое семейство. Типовой вид рода - Ulemosaurus svijagensis. Череп высокий, верхние края глазниц утолщены и бугристы, длина черепа свыше 40 см. Височная впадина узкая. Передние зубы (резцы) мощные, уменьшаются кзади. На зубах сильно развит уступ («пятка»), зубы могут плотно смыкаться. Сохраняются небольшие клыки, почти неотличимые от других зубов. Предполагается, что зубная система была приспособлена к перетиранию растительного материала, но перетирание осуществлялось передними зубами, а не задними, как у большинства растительноядных позвоночных. Улемозавр (и все прочие тапиноцефалы) мог питаться мертвыми гниющими стволами каламитов и других растений. Передние ноги длиннее и массивнее задних, голова высоко приподнята над землей (как и у всех тапиноцефалов). Плечевой пояс заметно массивнее, чем у мосхопса, плечевая кость длиннее. Общая длина животного могла быть до 4 метров. Описан (по фрагментам) еще один вид — Ulemosaurus gigas. Он известен по отдельному резцу из Ключевского рудника (Башкирия) и частичному черепу из местонахождения Озерки (Оренбургская область). Судя по всему, кроме более крупных размеров, отличался от типового вида более длинной мордой.
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие синапсиды, Пермь, терапсиды |
| Time | Event |
| 04:22 pm [industrialterro] [Link] |
Venjukovia Venjukovia (Amalitskii, 1922) is an extinct genus of non-mammalian synapsid from the Middle Permian of Russia. Венюковии (Venyukoviidae) — семейство примитивных аномодонтов среднепермской эпохи. Образуют надсемейство Venyukovioidea. Описаны два рода, которые могут быть синонимами. Венюковия (Venyukovia prima) описана В. П. Амалицким по нижним челюстям из Каргалинских рудников в Оренбуржье. Остатки собрал в 1908 году профессор Петроградского горного института П. Н. Венюков, в честь которого и был назван род. Владимир Амалицкий считал венюковию млекопитающим. Позднее, в 1935 году череп подобного животного был обнаружен в Ишеево. Изучение ишеевского черепа и челюстей венюковии позволило И. А. Ефремову отнести животное к терапсидам. Ефремов сближал венюковию с дейноцефалами. Позднейшие исследования позволили установить её принадлежность к самым примитивным аномодонтам. Для черепа из Ишеева обнаружили отличия в строении зубов и он был отнесён к особому роду улемика (Ulemica). Таким образом, от венюковии известны только обломки нижних челюстей. В 1960 году П. К. Чудинов описал из Очерского местонахождения череп мелкого аномодонта очерии (Otsheria netzvetajevi). Это ближайший родич венюковии, который может относиться к этому же роду. В целом находки этой группы очень редки. Также к семейству венюковий отнесена суминия, обнаруженная близ Котельнича. Венюковии — некрупные животные, длина черепа 10—12 см. Череп довольно высокий, короткий, полукруглый, глазницы огромные. Передние зубы долотовидные, напоминают резцы грызунов. Щёчные зубы широкие, сильно стёртые. Возможно, могли перерабатывать пищу во рту (раздавливать). Хоанная область очень глубокая. Теменной бугор высокий. Нижняя челюсть массивная. Скуловая дуга изогнутая, по очертаниям череп сходен с таковым у дицинодонтов. Вероятно, наземные растительноядные, питались относительно мягкой растительностью. Могли питаться также насекомыми.
Tags: Вымершие синапсиды, Пермь, терапсиды |









































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Previous Day | 2011/12/04 [Archive] |
Next Day |