[Recent Entries][Archive][Friends][User Info]
Below are the 20 most recent journal entries recorded in the "Сообщество, посвящённое ра" journal:| March 13th, 2016 | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 03:06 pm [industrialterro] [Link] |
Amurosaurus Амурозавр (Amurosaurus) — птицетазовый динозавр из подсемейства ламбеозаврины (Lambeosaurinae). Единственный известный вид — Amurosaurus riabinini. Амурозавр обитал на Земле в конце позднего мела, 74—65 миллионов лет назад. Как и большинство ламбеозаврин, амурозавр был двуногим травоядным ящером из семейства гадрозавриды — «утконосых динозавров» с полым гребнем на вершине головы. Сам гребень амурозавра не был найден. Окаменелые кости взрослых экземпляров очень редки, но по имеющимся останкам можно судить, что амурозавр был длиной от 6 до 8 метров. Российские палеонтологи Юрий Болотский и Сергей Курзанов описали и дали название этому динозавру в 1991 году. Название произошло от реки Амур и древнегреческого слова sauros («ящер»). Река Амур протекает на границе России и Китая, где и были найдены останки данного динозавра. Объем мозга амурозавра был больше, чем у большинства других травоядных динозавров. Относительно большой мозг указывает на сложное поведение. Возможно, это отчасти адаптация к стайному образу жизни. Останки амурозавра хранятся в Хабаровском Краеведческом музее имени Гродекова. В музее также имеется модель динозавра в натуральную величину. Russian paleontologists Yuri Bolotsky and Sergei Kurzanov first described and named this dinosaur in 1991. The generic name is derived from the Amur River and the Greek word sauros ("lizard"). The Amur (called Heilongjiang or "Black Dragon River" in Chinese) forms the border of Russia and China, and is near where this dinosaur's remains were found. There is one known species (A. riabinini), named in honor of the late Russian paleontologist Anatoly Riabinin, who conducted the first Russian expeditions to recover dinosaur remains in the Amur region in 1916 and 1917. All fossils of Amurosaurus have been recovered from a single bonebed locality, discovered in 1984 within the city limits of Blagoveschensk in the Amur Oblast of far eastern Russia. This bonebed is found in the Udurchukan Formation, the oldest geologic formation in the Tsagayan Group of far eastern Russia and northeastern China. This formation is thought to belong to the Maastrichtian stage of the Late Cretaceous Period, which was deposited 66 million years ago in the equivalent of the Lancian faunal stage of North America. The sediments were laid down in the floodplain of a river, which transported the fossils, but only a short distance, judging by the randomly assorted, disarticulated, but well-preserved bones within the bonebed, including fragile skull elements. Only a small section of the bonebed has been uncovered, but 90% of the remains found so far belong to lambeosaurines like Amurosaurus, mostly juveniles, with the rest belonging to other taxa, such as the hadrosaurine Kerberosaurus. Theropod teeth are also abundant, and there are many toothmarks on the bones, made by predators or scavengers. The holotype, or original specimen, consists of only a maxilla (upper jaw bone), and a dentary (lower jaw bone), both from the left side of the same individual. However, most of the other bones of the skull and skeleton have also been preserved in the bonebed, albeit of many different individuals. This other material was described more recently, making Amurosaurus the most abundant and completely known Russian dinosaur. Amurosaurus is characterized by many autapomorphies, or unique features, of the skull, as well as the sigmoidal shape of the ulna (a lower arm bone) when viewed from the front or side. Most other known lambeosaurines have hollow crests on the top of their skulls, and although the bones that would make up such a crest are unknown in this dinosaur, the bones of the roof of the skull are modified to support one, so it can be assumed that Amurosaurus was crested as well. As most of its features were described recently, as of early 2006, Amurosaurus has only been subjected to one cladistic analysis, which placed it as a basal member of the lambeosaurine subfamily of hadrosaurs, but more derived than either Tsintaosaurus or Jaxartosaurus. All known basal lambeosaurines come from Asia, which has led to the hypothesis that lambeosaurines originated there and then later dispersed across the Bering Strait to North America. Two derived groups, the parasaurolophins (Parasaurolophus, Charonosaurus) and lambeosaurins (Corythosaurus, Nipponosaurus, Lambeosaurus, etc.) evolved later. As members of both groups are found in North America and Asia (And one from Europe, which one is not solidly known), there must have been further dispersal after their evolution, although in which direction that dispersal occurred is still unclear. ( Read More ) Гадрозавриды, или гадрозавры (Hadrosauridae) — семейство так называемых «утконосых» динозавров, подотряд орнитопод, которое делится на два подсемейства — гадрозаврины и ламбеозаврины. Время существования — меловой период. Для семейства гадрозавров характерен гребень различных форм. В этом гребне находились носовые полости, которые, вероятно, служили для издавания громких звуков. Гадрозавры имели плоский беззубый, схожий с утиным, клюв, а в задней части челюсти находилось множество плоских зубов. Передвигались как на двух, так и на четырёх конечностях. Имели длинные задние и более короткие передние лапы. Длинный плоский хвост служил для равновесия. Гадрозавриды вели стадный образ жизни, строили гнёзда, охраняли кладку и вместе заботились о потомстве. Долгое время гадрозавров, из-за строения передних конечностей с перепонками между 4 и 5 пальцами и плоским хвостом, считали полуводными водорослеядными животными. Сейчас же семейство «утконосых динозавров» относят к наземным обитателям. В желудке хорошо сохранившегося эдмонтозавра было найдено большое количество хвои, а не водорослей. Также установлено, что на передних конечностях у гадрозаврид были копытца, а под хвостом имелся мясистый нарост, из-за которого плавание было бы просто невозможным. Останки гадрозавров обнаружены в Азии, Антарктиде, Европе, Северной и Южной Америке. Lambeosaurinae — группа птицетазовых динозавров из семейства гадрозавридов, характерной особенностью которых является костное образование на голове. Lambeosaurinae включают в себя две трибы — Parasaurolophini (Parasaurolophus) и Corythosaurini (Corythosaurus, Hypacrosaurus и Lambeosaurus). Corythosaurini и Parasaurolophini были введены в научную литературу в 2007 году. Corythosaurini определяются как все таксоны, более тесно связанные с Corythosaurus casuarius, чем с Parasaurolophus walkeri и Parasaurolophini — все таксоны, более близко связанные с P. walkeri, чем с C. casuarius. Ископаемые останки и реплики (1, 2, 3, 4, 5): Tags: Вымершие рептилии, Мел, авеметатарзалии, анкилополлексии, архозавроморфы, архозавры, гадрозавриды, гадрозавроиды, диапсиды, динозавроморфы, динозавры, игуанодонты, ламбеозаврины, неорнитискии, орнитоподы, птицетазовые, цераподы | ||||||||||||||||||||||||||||||||||||||||
| February 11th, 2016 | |||||||||||||||||||||||||||||||||||||||||
| 11:06 pm [industrialterro] [Link] |
Ampelosaurus Ampelosaurus (/ˌæmpᵻloʊˈsɔːrəs/ am-pi-lo-sawr-əs; meaning "vine lizard") is a titanosaurian sauropod dinosaur hailing from the Late Cretaceous Period of what is now Europe. Its type species is A. atacis, named by Le Loeuff in 1995. A possible unnamed species has givenAmpelosaurus an age reaching to the latest Cretaceous, from about 70 to 66 million years ago. Like most sauropods, it would have had a long neck and tail but it also carried armor in the form of osteoderms. Over 500 bones have been assigned to Ampelosaurus and all but the braincase (assigned to A. sp.) has been assigned to A. atacis. They are assigned to the same species because all the differences in the limb proportions have been linked to individual variation. A. atacis is known from a few, well-preserved teeth and some cranial material. A right scapula was discovered associated with a coracoid. The blade of the scapula, contrary to most titanosaurs, is triangular. The blade narrows at one end instead of showing an expansion like most other genera. A. atacis is also known from a pubis about 75 centimetres (30 in) long and an ilium. Aside from that, it is known from a partial forelimb, and many femora. Titanosaurians were a flourishing group of sauropod dinosaurs during Cretaceous times. The Spanish locality from the latest Cretaceous of “Lo Hueco” yielded a relatively well preserved, titanosaurian braincase, which shares a number of unique features with A. atacis from France. However, it appeared to differ from A. atacis in some traits also. The specimen has been provisionally identified as Ampelosaurus sp.. A. sp. is most likely a mature titanosaurian since the bones of the braincase have largely fused together. Ampelosaurus lived alongside many other animals. Over 8500 specimens have been found alongside it, including gastropods, bivalves, crocodiles, other sauropods, plants and invertebrates in the Villalba de la Sierra, Gres de Saint-Chinian, Marnes Rouges Inférieures and Gres de Labarre formations. Recent attention has made Ampelosaurus one of the most well-known dinosaurs known from France. Like most sauropods, Ampelosaurus would have had a long neck and tail, but it also carried armor in the form of osteoderms 25 to 28 centimetres (9.8 to 11.0 in) long. The four osteoderms found have three different morphologies, they are plate, bulb, and spine-shaped. This dinosaur would have stretched up to about 15 metres (49 ft) from snout to tail. Ampelosaurus atacis is known from many unattached bones and teeth. It is one of the best-known dinosaurs from France. It is considered to be a dwarf sauropod by Coria et al. (2005) because it has a smaller size in comparison with its ancestors. Ampelosaurus is one of the most completely known titanosaurian sauropod from Europe. Since 1989, over 500 bones discovered at the Upper Cretaceous Bellevue locality of southern France have been assigned to A. atacis. An abundant amount of well-preserved elements have been uncovered at Bellevue. In the original description, only a tooth and some vertebrae were examined. In 2005, a complete description was made by Le Loueff of all known material, allowing Ampelosaurus to be much better understood. All the material has been assigned to the species, and even though there are some differences in limb proportions, it has all been linked to individual variation. A. atacis is known from a few, well-preserved teeth and some cranial material, a tooth of which was described in its original description. The teeth differentiate Ampelosaurus from Magyarosaurus and Lirainosaurus, both of which are also from Europe. The teeth are different from the two later genera as Ampelosaurus has teeth that are roughly cylindrical in the top with thin expansions at the bottom. They are 21 millimetres (0.83 in) high and about 6 millimetres (0.24 in) wide. Those expansions give its teeth a slight constriction between the root and the crown. The teeth are also quite different from the peg-like teeth of titanosaurids. The teeth, have a general morphology similar to titanosauriansAntarctosaurus and a braincase from Dongargaon. Another skull was described from southern France, but showed some differences, meaning there could have been at least two different titanosaurians in France during the Late Cretaceous. The scapula was found attached to a coracoid. In the longer direction, the bone is 72 centimetres (28 in) long. The blade of the scapula, contrary to titanosaurs, is triangular, larger closer to the base. The blade narrows distally instead of showing an expansion. Differing fromMagyarosaurus and Saltasaurus, the scapula does not have the dorsal crest at the base of the scapular blade. The shape of the coracoid is quadrangular, and the coracoid has a thickened cranial margin. Ampelosaurus is also known from a pubis, about 75 centimetres (30 in) long. It has a strong distal expansion, that is similar to a titanosaur from Brazil, and a large foramen. Contrary to the South American titanosaurs, the ilium does not have an expansion on the side. The ischium, known from a shaft, is unexpanded distally and very well developed. It is also very flat. An incomplete radius is also known. It lacks the proximal and distal ends, and possesses a very prominent ridge along it. The largest and best-preserved specimens are both ulnas. The right ulna has a total length of 395 millimetres (15.6 in). The right ulna is small and slender, is missing an olecranon, and has a well-marked ridge. The left ulna is larger, with a length of 725 millimetres (28.5 in). The ulna has a deep radial fossa. About 27 femora are known from Bellevue, making them the most common bone, most of them more-or-less complete. They are very flat craniocaudally but otherwise, lack any unusual features. In a study of its bone physiology, it was found that the maximum known femur length was 1,100 millimetres (43 in), which is much larger than previously known (802 millimetres (31.6 in)). That means, that the ancestorsAmpelosaurus were slightly smaller than itself. 18 humeri are also known, but none of them approach the size of the largest femur. The humeri do not have a free medullary cavity. The humeri are robust, about 63 centimetres (25 in) long, with very expanded distal and proximal ends. Three different types of osteoderms have been recovered at Bellevue. Two with large spines have been uncovered. At the base of each spine, which is about 12 centimetres (4.7 in) tall, are two large foramina. The internal faces are all concavo-convex and have a ridge opposite the spine. In side view, they have two sections, a low half that progressively thickens, and a spine. ( Read More ) Размеры тела в сравнении с человеком: Ископаемые останки и реплики (1, 2, 3, 4, 5, 6, 7, 8): ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| June 25th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 05:08 pm [industrialterro] [Link] |
Amazonsaurus Amazonsaurus (/ˌæməzənˈsɔrəs/ AM-ə-zən-SAWR-əs, "Amazon lizard") is a genus of diplodocoid sauropod dinosaur from the Early Cretaceous Period of what is now South America. It would have been a large-bodied quadrupedal herbivore with a long neck and whiplash tail. Although more derived diplodocoids were some of the longest animals ever to exist, Amazonsaurus was probably not more than 12 meters (40 ft) long. Despite the fact that other dinosaurs have been found in Brazil, this is the first named genus from territory in the Amazon Basin. The generic name is derived from the Brazilian Legal Amazon region and the Greek word sauros ("lizard"). There is one named species, (A. maranhensis), which is named after the Brazilian state of Maranhão. Both genus and species were named in 2003 by Brazilian paleontologists Ismar de Souza Carvalho and Leonardo dos Santos Avilla, and their Argentine colleague, Leonardo Salgado. Fossils of Amazonsaurus, including some back and tail vertebrae, ribs, and fragments of the pelvis, are the only dinosaur remains identifiable at the generic level from the Itapecuru Formation of Maranhão. This geologic formation dates back to the Aptian through Albian epochs of the Early Cretaceous Period, or about 125 to 100 million years ago. Amazonsaurus was recovered in sediments which are interpreted by geologists as floodplain deposits near a river delta. The tall neural spines on the tail vertebrae identify Amazonsaurus as a diplodocoid sauropod, but the fragmentary nature of the only known specimen makes it difficult to place A. maranhensis more specifically within the superfamily Diplodocoidea. However, some features of these vertebrae suggest it may be a late-surviving member of a line of basal diplodocoids. At least one published cladistic analysis shows Amazonsaurus to be more derived than rebbachisaurids, but basal to dicraeosaurids and diplodocids within Diplodocoidea (Salgado et al., 2004). Basal diplodocoids are found in several parts of South America, as well as northern Africa, during the Early Cretaceous, as are titanosaurian sauropods, and the carcharodontosaurid and spinosaurid theropods. By the Late Cretaceous Period, the diplodocoids had gone extinct, while titanosaurs proliferated. The predatory theropod families of the Early Cretaceous were also replaced by abelisaurid theropods throughout the southern continents during the Late Cretaceous. Rebbachisauridae — семейство завроподовых динозавров из надсемейства Diplodocoidea, известных из фрагментарных останков мелового периода Южной Америки, Африки и Европы. Данная группа являются довольно плохо изученной среди завроподов. Типовой род семейства Rebbachisaurus, гигантский завропод найденный в Северной Африки, впервые был описан в 1954 году. С тех пор было описано несколько тесно связанных видов, в основном из Южной Америки. В 1990 году палеонтолог Джек Макинтош включил род Rebbachisaurus в семейство Diplodocidae, подсемейства Dicraeosaurinae на основе описанных останков скелета. С открытием в последующие годы дополнительных видов, стало ясно, что Rebbachisauridae составляют отдельную группу динозавров, а в 1997 году аргентинский палеонтологом Хосе Бонапарте было назначено новое семейство — Rebbachisauridae. Пол Серено и его сотрудники в 1999 году перегруппировали данное семейство. Несмотря на то, что представители семейства известны только из меловых отложений, они вероятно являются базальными членами надсемейства Diplodocoidea и, следовательно, более примитивны, чем ранние диплодоки и апатозавр, жившие в конце юрского периода, что указывает на большой пробел в эволюционном древе завропод от средней до поздней юры. Кроме Rebbachisaurus и Limaysaurus в семейство относят вид Nigersaurus taqueti, известный по нескольким хорошо сохранившимся скелетам из раннего мела (апт — альб) Нигера. Из десятка видов семейства до сих пор только Limaysaurus и Nigersaurus известны по относительно полным скелетам. Все другие останки очень фрагментарны, так что трудно получить полную картину для данного семейства. Одним из характерных общих признаков этой клады является своеобразная форма лопатки. Кроме Африки и Южной Америки Rebbachisauridae также обитали в Европе. Вид Histriasaurus boscarottii был описан на основе нескольких позвонков из раннего мела (готерив — баррем) Хорватии (Dalla Vecchia, 1998) и является самый ранним динозавром из этого семейства. Фрагменты костей из формации Кастрильо-де-ла-Рейна (баррем — апт) Испании в 2011 году были подробно описаны под именем Demandasaurus darwini. Также известны останки на острове Уайт в Англии. Фрагменты скелета (кость лопатки), найденные Ником Касе в 1989 году в формации Уэссекс на юго-западном побережье острова Уайт, были описаны Филипом Маннионом в 2009 году. В кладистическом анализе, проведённом Карбаллидо в соавторстве с другими учеными в 2010 году, полученные на основе сравнения лопаточных костей данные указывают на очень тесные связи английских реббахизаврид с южноамериканскими видами, что может свидетельствовать о том, что английский реббахизаврид может являться сестринским таксоном для южноамериканского вида Limaysausus tessonei. Вполне возможно, что описанный в 2011 году хвостовой позвонок принадлежат животному того же вида, несмотря на то что данные филогенетического анализа установили родство этого динозавра с испанским Demandasaurus и африканским Nigersaurus, а не с Limaysaurus. д Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, диплодоциды, завроподоморфы, завроподы, реббахизавриды, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| June 21st, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 08:13 pm [industrialterro] [Link] |
Amargasaurus Амаргаза́вр (Amargasaurus, букв. «ящер из каньона Ла-Амарга») — род динозавров, живших в Аргентине 130 млн лет назад. Название «амаргазавр» происходит от названия каньона (Ла-Амарга), где были найдены останки. Амаргазавр описан и назван Хосе Бонапарте в 1991 году. Точная длина неизвестна, так как хвост не был найден, однако она равнялась приблизительно 9-12 м (по некоторым источникам 14 м), высота — 3-3,7 м, вес — до 9000 кг. Отличительные черты этого вида: слегка выступающие из челюсти зубы и два ряда 65-сантиметровых шипов на верхней стороне тела. Согласно одной из теорий, шипы были покрыты роговой тканью и предназначались для защиты от хищников, а если верить другой теории, то у амаргазавра был некий парус, пронизанный кровеносными сосудами, как у спинозавра. Учитывая, то, что эта конструкция значительно ограничивала подвижность шеи амаргазавра нельзя сказать, что у ящера был парус; у динозавра не хватило бы сил, чтобы поднять всё это, да к тому же не смог бы поворачивать голову, однако у шипов были и плюсы; они помогали в брачных играх и защите от хищников. Научно описаны два вида амаргазавра: A. cazaui и A. groeben. Ареал амаргазавра ограничен территорией нынешней провинции Неукен (Аргентина). В Аргентине с готеривского по аптский отделы мелового периода с амаргoзавром обитали абелизаврид лигабуейно и завропод агустиния. Растительность тоже не отличалась разнообразием: в первую очередь это древовидные папоротники и хвойные (араукарии и сосны). Амаргазавр, по-видимому, питался араукариями и древовидными папоротниками.
Этот динозавр рос сравнительно медленно для зауропода.
Amargasaurus (/əˌmɑrɡəˈsɔrəs/; "La Amarga lizard") is a genus of dicraeosaurid sauropod dinosaur from the Early Cretaceous Period (129.4–122.46 mya) of what is now South America. It was small for a sauropod, reaching 10 meters (33 feet) length. It would have been a quadrupedal herbivore with a long, low skull on the end of a long neck, much like its relative Dicraeosaurus. However, this dinosaur sported two parallel rows of tall spines down its neck and back, taller than in any other known sauropod. These spines have been reconstructed supporting skin sails, but the "skin sail" hypothesis was rejected by Gregory S. Paul in 2000. It was found at the La Amarga Arroyo (which means "the bitter creek" in Spanish) in Neuquén Province, Argentina. They are related to Diplodocus, a giant, long-necked sauropod from the Late Jurassic of North America. However they went down a different evolutionary path to their ancestors and evolved a much shorter neck and a drastically reduced overall body size. Amargasaurus was small for a sauropod, measuring 9 to 10 meters in length and weighing approximately 2.6 tons. It moved on four legs and probably was unable to rear on its hind legs. The neck of Amargasaurus, measuring 2.4 meters in length, was proportionally short for sauropod standards. It consisted of 13 cervical vertebrae which were opisthocoelous (convex at the front and hollow at the back), forming ball-and-socket joints with neighbouring vertebrae. The trunk was made out of 9 dorsal and probably 5 fused sacral vertebrae. While the foremost dorsals were opisthocoelous, the remaining dorsals were amphyplatyan (flat on both ends). Robust transverse processes (lateral projections connecting to the ribs) indicate a strong developed rib cage. The dorsal vertebrae of Amargasaurus and other dicraeosaurids lack pleurocoels, the deep lateral excavations that were characteristic for other sauropods. The most obvious feature of the Amargasaurus' skeleton were the extremely tall, upwardly projecting neural spines on the neck and foremost back vertebrae. The neural spines were bifurcated along their entire length, forming a double row. They were circular in cross section and tapered towards their tips. The tallest spines could be found on the middle part of the neck, where they reached 60 cm on the 8th cervical. On the neck, they were bowed backwards, projecting above the adjacent vertebra. Greatly elongated spines continue along the last two dorsal vertebrae, the hip and foremost tail. However, in these regions the spines were not bifurcated but flared into a paddle-shaped upper end. The skull is only incompletely preserved––however, it likely had a horselike, broad snout equipped with pencil-like teeth, based on related sauropods for which more complete skulls are known. As in other dicraeosaurids, the external nares (nostril openings) were situated in the posterior half of the skull, diagonally above the eye sockets, which were proportionally large. Additional (fenestrae) could be found surrounding the eye sockets: The infratemporal fenestrae, located below the eye sockets, were long and narrow. Behind the eye sockets were the supratemporal feneatrae, which, in dicraeosaurids, were uniquely small and directed laterally (in contrast to other diapsids, where they were directed upwards). In front of the eyes would have been the antorbital fenestra, although this region is not preserved. Small openings, called parietal openings or fontanelles, can be seen on the backside of the skull. In other tetrapods, these openings are usually seen only in juveniles and they close as the individual grows. The name Amargasaurus was coined in 1991 by Argentine paleontologists Leonardo Salgado and José Bonaparte, because its fossil remains were found alongside the La Amarga Arroyo in the province of Neuquén in Argentina. La Amarga is also the name of a nearby town, as well as the geologic formation the remains were recovered from. The word amarga itself is Spanish for "bitter," while sauros is Greek for "lizard." The one named species (A. cazaui) is named in honor of the man who discovered the site, Luis Cazau, a geologist with the YPF oil company, which at the time was state-owned. This site is located in the lower (older) sections of the La Amarga Formation, which dates to the Barremian through early Aptian stages of the Early Cretaceous Period, or around 130 to 120 million years ago. Amargasaurus is known from a relatively complete skeleton from a single individual. This skeleton includes the back of the skull, and all vertebrae of the neck, back, and hips, as well as a bit of the tail. The right side of the shoulder girdle is also known, as are the left forelimb and hind limb, and the left ilium, a bone of the pelvis. The spines may have supported a pair of tall skin sails. Similar sails are seen in the unrelated dinosaurs Spinosaurus and Ouranosaurus, as well as the pelycosaurs Dimetrodon and Edaphosaurus. There are a variety of hypotheses for the function of these sails, including defense, communication (for mating purposes or for simple species recognition), or temperature regulation. Gregory Paul argued that parallel neck sails would have reduced neck flexion. Instead, he proposed that, with their circular rather than flat cross-sections, these spines were more likely covered with a horny sheath. He even suggests that they could have been clattered together for a sound display. Similar spines are found on the presacral vertebrae of Dicraeosaurus from Africa, although not nearly as tall. Amargasaurus and Dicraeosaurus are united with the more recently discovered Brachytrachelopan in the family Dicraeosauridae. Dicraeosaurids and diplodocid sauropods are included in a group called Flagellicaudata.
Амаргазавр - завропод раннего мелового периода был не очень крупным динозавром. В длину динозавр достигал всего десять метров и был гораздо короче своих гигантских родственников. Эта растительноядная рептилия была ростом до двух метров и весом до пяти тонн. Замечательной и главной особенностью амаргазавра являлись шипы длиной до шестидесяти пяти сантиметров, располагавшиеся на шее, спине и хвосте. Одного ряда шипов было бы вполне достаточно, чтобы динозавр имел странный вид, но двойной ряд - это просто ошеломляющее зрелище! Ряды шипов могли составлять особую гриву, а могли быть покрыты кожей и представлять собой подобие двойного паруса (гребня). Палеонтологи долгие годы спорили о том, какую же функцию могли выполнять эти шипы. Они расположены не слишком удобно, чтобы служить защитой от хищников, хотя не исключено, что амаргазавр мог ими отпугивать хищников, ведь удлиненные шипы были покрыты роговой оболочкой, и он выглядел со стороны как гигантский дикобраз. Может быть, они служили, чтобы животное казалось больше, чем есть на самом деле? Или этот гребень мог быть каркасом для своего рода паруса, выполнявшего роль терморегулятора в теле? Или же этот подарок эволюции играл особую роль в социальной жизни животного, помогал узнавать друг друга, добиваться определенного статуса в стаде и служить для брачных ритуалов? А может это был своеобразный горб, как у современного верблюда? Много вопросов, много теорий и предположений, но пока ученые не могут дать однозначного ответа, и роль шипов в жизни амаргазавра до сих пор остается под завесой тайны. До конца неясно, какую они могли также принимать форму, но, несомненно, это было очень нужное и уникальное приспособление. В любом случае эти шипы были хорошим преимуществом для ящера, который был практически вдвое короче своих сородичей. Дополнительные конструкции вроде этой побуждают ученых искать новые пути в палеонтологии. Такое впечатление, что эволюция берет простые и понятные части динозавров имеющих очевидные функции и наделяет их озадачивающими дополнениями, назначение которых загадочно и зачастую противоречиво. Шипастым позвоночником амаргазавр напоминал другого ящера – африканского зауропода дикреозавра (Dicraeosaurus) и поэтому некоторые палеонтологи выделяют эти два вида в особое семейство Dicraeosauridae. Палеонтологи связывают этих животных из-за наличия у них схожих шипов на пресакральных и ретросакральных позвонках, хотя у Dicraeosaurus эти шипы по высоте значительно ниже. Палеонтолог Грегори Пол утверждал, что двойной парус (шипы, гребень) имел свои недостатки: он ограничивали шею в движениях, уменьшая ее сгибание. Ученый даже предполагал, что амаргазавр мог стучать шипами друг о друга, издавая звуки при встрече. Шея амаргазавра состояла из тринадцати позвонков, которые были выпуклыми спереди и полыми сзади, образуя шаровое соединение (типа шарниров) с соседними позвонками. Шея образовывала сильную дугу, с очень высокой поднятой средней частью, и головой, как правило, всегда опущенной к самой земле. У амаргазавра была маленькая голова с длинным и низким черепом, у которого была суженная верхушка. Широкая выдающаяся вперед морда была оснащена тупыми карандашеподобными зубами, напоминавшими колышки. Такие зубы были отличным приспособлением для обрывания и перемалывания растительности. Как и у других дикреозаврид, внешние ноздри (отверстия ноздри) были расположены в задней половине черепа, по диагонали над глазницами, которые были пропорционально большими. Подвисочные отверстия, расположенные ниже глазниц, были длинные и узкие. За глазницами были надвисочные отверстия, которые у дикреозаврид были маленького размера и направлены в стороны (в отличие от других диапсидных рептилий, у которых они были направлены вверх). В передней части глаза были подглазничные отверстия, хотя эта область не сохранилась. Малые отверстия, называемые теменными отверстиями или родничками, располагались на задней стороне черепа. У других тетрапод эти отверстия обычно наблюдаются только у молодых особей, и они закрываются по мере роста. Амаргазавр передвигался на четырех мощных конечностях. Передние конечности были более короткими, чем задние. На всех его колоннообразных ногах было по пять пальцев. Один из пальцев на каждой из конечностей имел коготь, возможно использовавшийся для защиты. Как предполагают ученые, возможно, амаргазавры могли вставать на задние лапы, чтобы обрывать листву с верхушек деревьев или отпугивать врагов. Также для защиты динозавр использовал свой тонкий хлыстообразный хвост. В остальном же этот завропод мало отличался от своих сородичей.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10): ( Read More ) Сравнение размеров тела с человеком и другими дикреозавридами: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, дикреозавры, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| April 29th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 08:35 pm [industrialterro] [Link] |
Alxasaurus Alxasaurus elesitaiensis — вид динозавров-тероподов, принадлежащих к группе Maniraptora, живших в течение раннего мела в районе современного Китая. Голотип, IVPP 88402a, был найден в формации Bayin Gobi, которая восходит к апту. Он состоит из частичного скелета с нижней челюстью. Включает в себя большую часть позвоночника, передние конечности, части таза, бедра и задние кости. Также есть и другие находки: IVPP 88301, 88402b IVPP, IVPP 88501 и 88510 IVPP. В 2003 году был найден частичный череп Alxasaurus, который пока не получил инвентарный номер. Alxasaurus представляет собой теризинозавра среднего размера. Рассел и Донг в 1994 году дали оценку высоты динозавра в 3,8 метра. Грегори С. Пол в 2010 году оценил длину в 4 метра и вес в 400 килограммов. Длина экземпляра IVPP 88402b оценивается в 249 дюймов, вес в 110 килограммов, длина экземпляра IVPP 88501 оценивается в 283 дюйма, а вес в 160 килограммов. Alxasaurus был помещен в семейство Alxasauridae, другие же исследования относят его к кладе Therizinosauroidea. Alxasaurus (/ˌɑːlʃəˈsɔrəs/; "Alxa Desert lizard") is a genus of therizinosauroid alxasaurid theropod dinosaur from the Early Cretaceous Period of Inner Mongolia. It is one of the earliest known members of the superfamily Therizinosauroidea, but it already possessed the body shape - including the long neck, short tail, and long hand claws - of later therizinosauroids. Like other members of this group, it was a bipedal herbivore with a large gut to process plant material. Several specimens are known and the largest was a little over 12 feet (3.8 m) long. This dinosaur was first described and named by Canadian paleontologist Dale Russell and his Chinese colleague, Dong Zhiming, in a paper published in 1993. However, although the paper is technically included in the last issue of the 1993 volume of the Canadian Journal of Earth Sciences, this issue was actually released in the early weeks of 1994. Alxasaurus is named after the Alxa Desert of Inner Mongolia, also known as the "Alashan" desert, and the name also includes the Greek word sauros ("lizard"). Alxa (or Alashan) is also the name of the league, or administrative division, of the Inner Mongolia (Nei Mongol Zizhiqu) region of the People's Republic of China. The single known species (A. elesitaiensis) is named after Elesitai, a village found in this region, near which the fossil remains of Alxasaurus were located. Five Alxasaurus skeletons were recovered from the Bayin-Gobi Formation of Inner Mongolia, which dates to the Albian stage of the Early Cretaceous Period, or about 112 to 100 million years ago. The holotype, specimen IVPP 88402a, which is considered to exemplify the genus and species, is the largest and most complete of the five, consisting of the mandible (lower jaw) and some teeth, as well as many limb bones, ribs, and vertebrae, including all five sacral (hip) vertebrae and the first nineteen tail vertebrae. The other four specimens are the paratypes: IVPP 88301, IVPP 88402b, IVPP 88501 and IVPP 88510. Together the skeletons represent most of the bones in the body aside from the skull. While exhibiting many typical therizinosaur features in overall body shape and in the teeth, the skeleton of Alxasaurus also shows several features present in more typical theropods, and the discovery of this animal provided significant evidence that therizinosaurs were aberrant theropods. Specifically, the semilunate carpal bone of the wrist is found only in maniraptoran theropods, which also include oviraptorosaurs, dromaeosaurs, troodontids, and birds. Even more basal therizinosaurs such as Falcarius and the feathered Beipiaosaurus have since been discovered with more theropod features and have helped to solidify this arrangement. Alxasaurus is now thought to occupy a position between the early Beipiaosaurus and later therizinosaurs such as Erlikosaurus, Segnosaurus, and Therizinosaurus itself. Теризинозавры (или Сегнозавры) — инфраотряд тероподовых динозавров, относящийся к манирапторам (Maniraptora), который включает в себя одно надсемейство. Окаменелости теризинозавров были найдены в отложениях возрастом от раннего до позднего мела на территории Монголии, Китая и западной части США. Различные особенности передних конечностей, черепа и таза позволяют объединить их с тероподами манирапторами — близкими родственниками птиц, несмотря на внешние различия. Название «теризинозавры» происходит от названия одного из представителей этой группы — теризинозавра. Название «сегнозавры» происходит от названия другого представителя этой группы — сегнозавра. ( Read More ) Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, манирапторы, теризинозавры, тероподы, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| 07:26 pm [industrialterro] [Link] |
Alvarezsaurus Альваресзавр (от Alvarezsaurus — «ящер Альвареса») — род динозавров, живший 88—74 млн лет назад (с коньякского по сантонский ярусы позднего мелового периода) на территории нынешней провинции Неукен (Аргентина). Достигал длины 2 метра, высоты 0,7 метра и веса 20 кг. Впервые описали Х. Бонапарте в 1991 году. У альваресзавра отсутствуют шипы на спинных позвонках, в результате чего тело выглядит компактным. Отсутствие гребня, тянущегося вдоль спины, доказывает, что он был очень похож на птицу. Хвост, уплощённый с боков, очень длинный, примерно вдвое длиннее тела и шеи. Шея длинная и гибкая. У альваресзавра были длинные, лёгкие ноги, типичные для бегающего животного. Он считается самым примитивным представителем группы альваресзаврид. Скорее всего, эти ящеры питались насекомыми, мелкими земноводными, возможно, падалью и яйцами других рептилий. Строение их задних конечностей позволяет предполагать, что они неплохо лазали по поваленным и наклонённым стволам деревьев в поисках пищи. Череп и передние конечности альваресзавра (крайне необходимые для установления принадлежности к группе и реконструкции внешнего облика динозавра) не были обнаружены, и только после того как были найдены останки других членов клады, учёные смогли оценить, насколько необычными были эти динозавры. Первоначально альваресзавра реставрировали с типичными трёхпалыми конечностями целурозавра, в то время как, на самом деле, передние конечности альваресзаврид несли на себе по одному единственному пальцу. Alvarezsaurus (/ˌælvərɛzˈsɔrəs/; "Alvarez's lizard") is a genus of small alvarezsaurid dinosaur from the Late Cretaceous period of Argentina, approximately 86 - 83 million years ago. Estimates suggest that it measured about 2 meters (6.5 ft) in length and weighed approximately 20 kg (45 lbs). It was found in the Bajo de la Carpa Formation and was named by paleontologist José Bonaparte in 1991 after the historian Don Gregorio Alvarez. The type species is A. calvoi. It was bipedal, had a long tail, and its leg structure suggests that it was a fast runner. It may have been insectivorous and was basal to better-known members of its family, such as Mononykus and Shuvuuia. It has been alternately classified with both non-avian theropod dinosaurs and early birds.
Сравнение размеров нескольких альваресзаврид с человеком (альваресзавр под № 1): Tags: Вымершие рептилии, Мел, авеметатарзалии, альваресзавриды, альваресзавры, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, манирапторы, тероподы, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| April 25th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 12:48 pm [industrialterro] [Link] |
Aletopelta Алетопельта (Aletopelta) — род птицетазовых динозавров из семейства анкилозаврид, живших в кампанском веке мелового периода (около 75—70 млн лет назад) в Северной Америке. Окаменелые останки были обнаружены в Южной Калифорнии, США. Типовой вид — Aletopelta coombsi. Название состоит из греческих слов aletes и pelte, что означает, соответственно, «блуждающий» и «щит». Такое название этому роду динозавров было предложено Беном Крейслером, поскольку местность, где жил этот динозавр, в те времена находилась на гористом полуострове напротив современной Мексики. Тектоническая плита с полуостровом постепенно двигалась на север, неся останки динозавра с собой. Видовое название дано в честь Вальтера П. Кумбса-младшего за его новаторскую работу по анкилозаврам и многолетние исследования, которые вдохновляли профессиональных палеонтологов и простых энтузиастов. Интересна история голотипа алетопельты. Эта особь утонула в реке, течение вынесло труп в море, там он затонул окончательно и вокруг панциря вырос коралловый риф. Палеонтологи, откопавшие этот экземпляр, были сильно удивлены. Aletopelta coombsi /əˌliːtɵˈpɛltə ˈkoʊmzi/ is an herbivorous ankylosaurian ornithischian dinosaur that during the Late Cretaceous lived in the area of what is now Southern California. In 1987, construction work was done on the College Boulevard near Carlsbad at the Californian coast. In the Letterbox Canyon between El Camino Real and Palomar Airport Road, northwest of McClellan–Palomar Airport, the road was widened. While paleontologically surveying the work, Bradford Riney noted that a skeleton had been uncovered by a ditch dug for a sewage pipe. Within days, it was secured by the San Diego Natural History Museum. In 1988, the find was reported by Thomas Deméré. The discovery drew much attention because it was the first important ankylosaurian fossil known from the area. It was dubbed the "Carlsbad Ankylosaur". In 1996, the fossil was described but not named by Deméré and Walter Preston Coombs. In 2001, the type species Aletopelta coombsi was named by Tracy Lee Ford and James Kirkland. Etymologically, the generic name is composed of the Greek terms ἀλήτης, aletes, and πέλτη, pelte, meaning respectively "wanderer" and "small shield". This genus name was suggested by Ben Creisler because the fossil location, at the time the dinosaur died, being located on the tectonic plate containing the Peninsular Ranges Terrane, was somewhere opposite the middle of Mexico. This plate had thus been wandering northward since, carrying the specimen with it. The specific epithet honors the vertebrate paleontologist Walter Preston Coombs, Jr., "for his ground-breaking work on ankylosaurs and his years of research, which have inspired many an enthusiast as well as professional paleontologist". Aletopelta is known from a single partial skeleton lacking the skull, holotype SDNHM 33909, part of the collection of the San Diego Natural History Museum, San Diego, California. The skeleton including femora (the thighbones), tibiae (shinbones), fibulae (calf bones) and incomplete parts of a scapula (shoulder blade), humerus, ulna, left and right ischium, vertebrae, ribs, partial armor over the pelvic girdle, a cervival halfring plus at least sixty detached armor plates and eight teeth was found in a layer of the Late Cretaceous (Upper Campanian) marine Point Loma Formation, dating from the late Campanian, in 2001 estimated at 75.5 miliion years old. Apparently, the animal's bloated carcass floated out to sea, and formed a miniature reef environment after it sunk to the bottom, landing on its back, as testified by Pelecypoda attaching to the bones. The remains were scavenged by sharks. Most long bones have lost their joint surfaces and were hollowed out. As a result the condition of the skeletal elements is poor. Aletopelta is a medium-sized ankylosaurid, originally estimated to be around 6 metres (20 ft) long. In 2010, Gregory S. Paul estimated its length at five metres, its weight at two tonnes. Due to the damaged state of the holotype bones, it is difficult to determine distinguishing traits. In 2004, Matthew Vickaryous e.a. therefore considered Aletopelta to be a nomen dubium. Aletopelta is diagnosed as an ankylosaurid mainly based on the shape and arrangement of its osteoderm armor, which is closer in form to ankylosaurids than to nodosaurids. Анкилозавриды (Ankylosauridae) — семейство бронированных динозавров живших в меловом периоде 125 - 65 миллионов лет назад (вместе с другим семейством анкилозавров — нодозавровыми). Анкилозавриды были обнаружены в западной части Северной Америки, Европы и Восточной Азии, хотя хорошо сохранившиеся останки редки; наиболее известные находки состояли только из отдельных костных фрагментов. Анкилозавриды имели очень широкое, приземистое тело. На спине имелась броня, состоявшая из поперечных полос, составленных костными пластинами и узелками. Эти пластины соединялись отдельными кожными участками, благодаря чему шея, туловище и хвост сохраняли подвижность. Подобное строение брони имеют современные броненосцы. У многих анкилозавровид на шее и плечах были расположены длинные шипы, которые защищали животное от боковой атаки. У представителей подсемейства анкилозаврин на хвостах имелось мощное костяное утолщение, с помощью которого животные защищались от хищников. Они могли орудовать этим утолщением как средневековые рыцари булавой, благодаря чему динозавры и получили своё название. Возможно, эта булава была ярко раскрашена, на ней могли быть два ярких пятна, похожих на глаза, чтобы ещё больше сбить хищника с толку. У некоторых анкилозаврин эта булава могла весить более 27 килограммов. Тяжёлый панцирь и хвосты с роговыми частями делали анкилозаврин внешне похожими на млекопитающих глиптодонтов (и, в меньшей степени, на гигантских черепах миоланидов из Австралии). Анкилозавриды имели беззубый клюв в передней части челюстей, а в глубине рта у них имелись многочисленные мелкие зубы, характерные для тиреофор. Кладистический анализ, проведённый в 2011 году Томпсоном и его коллегами предполагает, что Minmi является наиболее базальным анкилозавром. Возраст данного динозавра оценивается примерно в 119—113 млн лет. На протяжении XX века верхний мел бассейна Сан-Хуан, штат Нью-Мексико, дал значительные образцы различных вымерших позвоночных, в том числе и анкилозаврида Nodocephalosaurus kirtlandensis. Находки анкилозавридов до начала XXI века в основном состояли из изолированных остеодермов и позвонков, известных из отложений в формациях Fruitland и Kirtland штата Нью-Мексико. В настоящее время Nodocephalosaurus kirtlandensis известен исключительно из формации Киртланд на основе частичного черепа и изолированных посткраниальных остеодерм. В 2005 году в данном регионе был найден новый вид анкилозавров — Ahshislepelta. Размеры тела в сравнении с человеком: Из останков торчит зуб акулы скваликоракса. Tags: Вымершие рептилии, Мел, авеметатарзалии, анкилозавриды, анкилозавры, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, птицетазовые, тиреофоры | ||||||||||||||||||||||||||||||||||||||||
| April 24th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 04:28 pm [industrialterro] [Link] |
Altirhinus Альтири́н (Altirhinus) — род птицетазовых динозавров из семейства игуанодонтид. Все окаменелости этого динозавра были найдены в 1981 году совместной Советско-Монгольской экспедицией в геологической формации Khukhtek, на территории Восточно-Гобийского аймака. Khukhtek датируется 125—100 миллионами лет, что относится к отложениям нижнего мела альбского—аптского ярусов. Известно несколько окаменелых скелетов. Типовым экземпляром является хорошо сохранившийся череп, а также некоторый посткраниальный материал, состоящий из частичных останков конечностей, плечевого и тазового поясов. Второй экземпляр включает частичный череп, связанный с рёбрами, фрагментарными позвонками, а также полными передними конечностями. Третий образец состоит из многих костей конечностей и 34 хвостовых позвонков оставшихся от меньшей особи. Невдалеке были обнаружены два скелета, вероятно, принадлежащим молодым особям этого вида. Первоначально останки альтирина были отнесены к виду Iguanodon orientalis, который был выделен в 1952 году. Однако, как показали дальнейшие исследования название Iguanodon orientalis было основано на чрезвычайно фрагментарных останках, которые вероятно на самом деле принадлежали Iguanodon bernissartensis. Останки, обнаруженные в Монголии, не соответствовали останкам игуанодона, поэтому в 1998 году они были выделены в отдельный род и вид. Altirhinus was herbivorous and bipedal when walking or running, but probably became quadrupedal when feeding from the ground. According to the original description, the entire body probably extended 26 feet (8 m) from snout to tail tip. In 2010 Gregory S. Paul estimated the length at 6.5 metres (21 ft), the weight at 1.1 tonnes. The skull alone is about 30 inches (760 mm) long, with a wide mouth and a distinctive tall arch on top of its snout, from which this dinosaur derives its name. All known specimens of Altirhinus were recovered in 1981 during collaborative expeditions organized by Soviet and Mongolian scientists, from the Khukhtek Formation in the Dornogovi Province of Mongolia. The Khukhtek was formed in the Aptian to Albian stages of the Early Cretaceous Period, which lasted from between 125 and 100 million years ago. Psittacosaurus and the primitive ankylosaurid Shamosaurus have also been found in these rocks. Several fossil specimens of different ages and sizes are known. The holotype, PIN 3386/8, is a skull, which is well preserved on the left side, as well as some postcranial material consisting of pieces of the hands, feet, shoulder and pelvic girdles. A more fragmentary skull was also recovered, associated with some ribs, fragmentary vertebrae, and a complete forelimb. A third specimen preserves many limb bones and a series of 34 tail vertebrae from a smaller individual. Two even smaller fragmentary skeletons, presumably of young individuals, were uncovered nearby. The remains of this animal were originally referred to the species Iguanodon orientalis, which was first described in 1952. However, I. orientalis has since been shown to be fragmentary, nondiagnostic, and virtually indistinguishable from the European I. bernissartensis (Norman, 1996). As no features of I. orientalis are shared exclusively with the 1981 specimens, which are clearly distinguishable from Iguanodon, a new name for those specimens was required. British paleontologist David B. Norman named them Altirhinus kurzanovi in 1998. The name was created from a Latin word, altus ("high") and a Greek word, ῥίς, rhis, genitive rhinos ("nose" or "snout"). There is one known species (A. kurzanovi), which honors Sergei Kurzanov, the influential Russian paleontologist who originally found the specimens in 1981. Altirhinus is definitely an advanced iguanodontian, just basal to the family Hadrosauridae, but there is little agreement on the arrangement of genera and species in this area of the ornithopod family tree. In the original description, it was included with Iguanodon and Ouranosaurus in a family Iguanodontidae (Horman, 1998). More recent analyses all find Altirhinus more derived than either of those two genera, but less than Protohadros, Probactrosaurus, and hadrosaurids (Head, 2001; Kobayashi & Azuma, 2003; Norman, 2004). The former two studies also place Eolambia between Altirhinus and hadrosaurids, while Norman's analysis finds that the two genera share a clade. Fukuisaurus is just basal to Altirhinus according to the only analysis in which the former has been included (Kobayashi & Azuma, 2003). Many aspects of Altirhinus' anatomy allow speculation on its behavior. As its forelimbs were roughly half the length of its hindlimbs, Altirhinus appears to have been primarily bipedal. However, its carpals (wrist bones) were thick and blocky, and the three middle fingers of its hand were wide, hyperextendable, and ended in hoof-like bones. This indicates that the forelimbs were also capable of supporting weight. Like many ornithopods, Altirhinus may have spent a significant amount of time in a quadrupedal position, perhaps while feeding. While the three middle digits of each forelimb (digits II, III, & IV) were very thick and probably weight bearing, the outside fingers (digits I & V) were modified in different ways. The first digit was a simple sharp spike, as seen in Iguanodon. Aside from defense, the thumb spike could possibly have also been used for breaking the shells of seeds or fruit. The fifth digit was somewhat opposable to the rest of the hand and may have been useful for grasping food. There is a large diastema, or gap, between the keratinous beak on the front of the mouth and the main chewing teeth in the side of the mouth, which would allow the two sections to work independently, so Altirhinus could crop with its beak while simultaneously chewing with its teeth. Many herbivorous mammals show a similar adaptation and can crop with their incisors without disturbing their chewing molars. Altirhinus was one of a number of advanced iguanodontians with snouts expanded outwards towards the end. This is quite possibly an example of convergent evolution with hadrosaurids, famous for their wide "duckbill" snouts. These adaptations are also paralleled in many living mammalian herbivores of different lineages. Modern cows, horses, and white rhinoceros all exhibit wide muzzles and all are grazing animals. Grazing most often occurs at ground level, and if the expanded muzzles of Altirhinus and other related species were an adaptation to grazing, this may also explain corresponding weight-bearing adaptations of the forelimbs in derived iguanodontians, in order to get the head closer to the ground. The characteristic arched snout of Altirhinus was formed primarily by the nasal bones, and a similar structure is seen on the snout of the Australian Muttaburrasaurus. Many different functions have been proposed for the nasal arch. It may have housed tissues to cool the blood, conserve water, or enhance the sense of smell. Alternatively, it may have facilitated communication through vocalization or visual display. As only two skulls have been located, it is entirely possible that the arched snout is only found in one gender, in which case it may have been used for sexual display, like in modern-day elephant seals. Гадрозавроиды (Hadrosauroidea) — клада или надсемейство птицетазовых динозавров, который включает в себя «утконосых» динозавров, или гадрозавридов, а также их близких родственников. Жили в меловом периоде на всех континентах планеты. Предполагается, что гадрозавроиды возникли в Азии и а потом широко расселились в другие части света. Самые ранние находки представителей данной группы относятся к аптскому веку раннего мелового периода, около 125 млн лет назад — это Probactrosaurus, живший на территории Китая, и Siamodon nimngami, найденный в Таиланде. Впервые название таксону дано Полом Серено в 1986 году в качестве подтаксона Iguanodontoidea. Ныне данная классификация не используется, и разные ученые предположительно относят гадрозавроидов либо к группе Hadrosauriformes (например Пол Серено в 1999 году и Годефруа (Godefroit) с коллегами в 2004 году), либо к группе Iguanodontia (например Ruiz-Omeñaca в 2006 году). Многие примитивные гадрозавроиды, такие как Ouranosaurus, традиционно включались в парафилетические группы игуанодонтидов. После кладистического анализа традиционных игуанодонтидов, данная группа в значительной степени распалась, и на данный момент включает в себя только игуанодона и его ближайших родственников. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, анкилополлексии, архозавроморфы, архозавры, гадрозавроиды, диапсиды, динозавроморфы, динозавры, игуанодонты, неорнитискии, орнитоподы, птицетазовые, цераподы | ||||||||||||||||||||||||||||||||||||||||
| 01:26 pm [industrialterro] [Link] |
Alioramus Алиорам (Alioramus, от лат. alius ramus «другая ветвь») — род хищных динозавров из семейства тираннозавридов, живших в меловом периоде (около 70—65 млн лет назад) на территории современной пустыни Гоби (Монголия). Известны два вида. Впервые алиорама (A. remotus) описал советский палеонтолог С. М. Курзанов в 1976 году, обнаружив в монгольских отложениях фрагменты черепа и три плюсневые кости, принадлежащие динозавру, который жил 70-65 млн лет назад. Название от лат. alius ramus «другая ветвь» было выбрано из-за анатомических отличий алиорама от других тираннозаврид. С тех пор вопрос о принадлежности алиорама к тираннозавридам оставался дискуссионным. Новые останки обнаружены в монгольской пустыне Гоби в 2001 году сотрудниками Американского музея естественной истории под руководством Марка Норелла и Майкла Новачека. Они, вероятно, принадлежат девятилетнему подростку, который вырос всего на 85 %, жил 85—65 млн лет назад, достигал в длину 5 метров и весил около 369 кг. В 2009 году данные останки окончательно отнесли к новому виду — Alioramus altai. Главное отличие алтайского алиорама — необычные рога. Помимо обычных для тираннозавридов надбровных коротких рогов у него имеется по два пятисантиметровых рога, торчащих из каждой щеки. Использовать их для борьбы невозможно, поэтому предполагается, что рога служили украшением для привлечения самок. Алиорам по сравнению тарбозавром обладал маленьким черепом, миниатюрными зубками и слабыми челюстными мышцами и занимал отличную от последнего экологическую нишу. Alioramus (/ˌæli.ɵˈreɪməs/; meaning 'different branch') is a genus of tyrannosaurid theropod dinosaurs from the Late Cretaceous period of Asia. The type species, A. remotus, is known from a partial skull and three foot bones recovered from Mongolian sediments which were deposited in a humid floodplain about 70 million years ago. These remains were named and described by Soviet paleontologist Sergei Kurzanov in 1976. A second species, A. altai, known from a much more complete skeleton, was named and described by Stephen L. Brusatte and colleagues in 2009. Its relationships to other tyrannosaurid genera are unclear, with some evidence supporting a hypothesis that Alioramus is closely related to the contemporary species Tarbosaurus bataar. Alioramus were bipedal like all known theropods, and their sharp teeth indicate that they were carnivores. Known specimens were smaller than other tyrannosaurids like Tarbosaurus bataar and Tyrannosaurus rex, but their adult size is difficult to estimate since both species are known only from juvenile or sub-adult remains. The recent discovery of Qianzhousaurus indicates that it belongs to a distinct branch of tyrannosaur. The genus Alioramus is characterized by a row of five bony crests along the top of the snout, a greater number of teeth than any other genus of tyrannosaurid, and a lower skull than other tyrannosaurids. Alioramus remotus was estimated at 5 to 6 metres (16 to 20 ft) in length when originally described by Sergei Kurzanov in 1976. Kurzanov, however, did not correct for lengthening of the skull by deformation during fossilization, which may indicate a shorter overall body length for this individual. If this specimen is a juvenile, then adult Alioramus would have reached greater lengths, but no confirmed adult specimens are known. The skull of A. remotus was approximately 45 centimetres (18 in) long. In general, it is long and low, a shape typical of more basal tyrannosauroids and juveniles of larger tyrannosaurids. The premaxillary bones at the tip of the snout in Alioramus remotus have not been found, but are taller than wide in all tyrannosauroids for which they are known. The nasal bones are fused and ornamented with a row of five irregular bony crests that protrude upwards from the midline, where the nasal bones are sutured together. These crests all measure more than 1 centimetre (0.39 in) tall. At the back of the skull there is a protrusion, called the nuchal crest, arising from the fused parietal bones, a feature shared with all tyrannosaurids. In Alioramus, the nuchal crest is greatly thickened, similarly to Tarbosaurus and Tyrannosaurus. Like the rest of the skull, the lower jaw of Alioramus was long and slender, another possible juvenile characteristic. As in Tarbosaurus, a ridge on the outer surface of the angular bone of the lower jaw articulated with the rear of the dentary bone, locking the two bones together and removing much of the flexibility seen in other tyrannosaurids. Other tyrannosaurids had four premaxillary teeth, D-shaped in cross section, on each side. Including 16 or 17 in each maxilla, and 18 in each dentary, Alioramus had 76 or 78 teeth, more than any other tyrannosaurid. The braincase of A. altai was intermediate between the basal theropod and avialan conditions. The rest of the skeleton of Alioramus remotus is completely unknown except for three metatarsals (bones of the upper foot), but the discovery of A. altai, which is known from substantially more complete remains, has shed light on the anatomy of the genus. Paleontologists have long classified Alioramus within the superfamily Tyrannosauroidea, but because its remains were for many years poorly known, a more precise classification had remained elusive until the discovery of A. altai. A cladistic analysis published in 2003 found Alioramus could be further classified into the family Tyrannosauridae and the subfamily Tyrannosaurinae, alongside Tyrannosaurus, Tarbosaurus and Daspletosaurus. A 2004 study supported this result but suggested it was equally probable that Alioramus belonged outside the family Tyrannosauridae entirely, with its supposed juvenile characters actually reflecting a more basal position within Tyrannosauroidea. Another study omitted Alioramus altogether due to the only specimen's fragmentary nature. The discovery of A. altai in 2009 confirmed the placement of the genus within the Tyrannosaurinae. Tarbosaurus and Alioramus shared several skull features, including a locking mechanism in the lower jaw between the dentary and angular bones, and both lacked the prong of the nasal bones which connected to the lacrimal bones in all other tyrannosaurids except adult Daspletosaurus. The two genera may be closely related, representing an Asian branch of the Tyrannosauridae. Some specimens of Tarbosaurus have a row of bumps on the nasal bones like those of Alioramus, although much lower. The long and low shape of the only known Alioramus remotus skull indicated that it was immature when it died and might even have been a juvenile Tarbosaurus, which lived in the same time and place. The more prominent nasal crests and much higher tooth count of Alioramus, however, suggested it was a separate taxon, even if it is known only from juvenile remains, confirmed by the discovery of A. altai. Specimens identified as immature Tarbosaurus have the same tooth count as adults. The holotype (PIN 3141/1) of Alioramus is a partial skull associated with three metatarsals. A joint Soviet-Mongolian expedition to the Gobi Desert in the early 1970s found these remains at a locality known as Nogon-Tsav in the Mongolian province of Bayankhongor. Alioramus was named and described by Russian paleontologist Sergei Kurzanov in 1976. Its crests and low skull profile looked so different from other tyrannosaurids that Kurzanov believed his find was far removed from other members of the family. Accordingly, he gave it the generic name Alioramus, derived from the Latin alius ('other') and ramus ('branch'), and the specific name A. remotus, which means 'removed' in Latin. Alioramus is known from the holotypes of A. remotus and A. altai. The Beds of Nogon-Tsav are considered to be the same age as the Nemegt Formation. This geologic formation has never been dated radiometrically, but the fauna present in the fossil record indicate it was probably deposited during the Maastrichtian stage, at the end of the Late Cretaceous. The Maastrichtian stage in Mongolia, as preserved in the Nemegt Formation and at Nogon-Tsav, was characterized by a wetter and more humid climate compared with the semi-arid environment preserved in the earlier, underlying Barun Goyot and Djadochta Formations. Nemegt sediments preserve floodplains, large river channels and soil deposits, but caliche deposits indicate periodic droughts. This environment supported a more diverse and generally larger dinosaur fauna than in earlier times. Kurzanov reported that other theropods, including Tarbosaurus, ornithomimosaurs and therizinosaurs were discovered at the same locality, but these remains have never been reported in detail. If the Nogon Tsav fauna was similar to that of the Nemegt Formation, troodontid theropods, as well as pachycephalosaurs, ankylosaurids and hadrosaurs would also have been present. Titanosaurian sauropods were also potential prey for predators in the Nemegt. ( Read More ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9): ( Read More ) Размеры тела в сравнении с человеком:
Ископаемые останки и реплики (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| April 21st, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 03:50 pm [industrialterro] [Link] |
Alectrosaurus Alectrosaurus (/əˌlɛktrɵˈsɔrəs/; meaning "alone lizard") is an extinct genus of tyrannosaurid theropod dinosaur that lived approximately 83 to 74 million years ago during the latter part of the Cretaceous Period in what is now Inner Mongolia. It was a medium sized, moderately-built, ground-dwelling, bipedal carnivore, with a body shape similar to its much larger relative, Tyrannosaurus rex, and could grow up to an estimated 5 m (16.4 ft) long. The generic name Alectrosaurus can be translated as "alone lizard," and is derived from the Greek words alektros and sauros ("lizard"). The meaning of the generic name is troublesome because the Greek work "alektros" (αλεκτρος) has also been alternatively translated as "unmarried" and even "rooster". At the time of its discovery, it was unlike any other Asian carnivore known, which may suggest that Gilmore selected a Greek word that reflected the meaning of alone. There is one named species (A. olseni), which is named in honor of George Olsen, who discovered the first specimens. Both genus and species were described and named by American paleontologist Charles Gilmore in 1933. In 1923, the Third Asiatic Expedition of the American Museum of Natural History, led by chief paleontologist Walter W. Granger was hunting for dinosaur fossils in Mongolia. On April 25th, assistant paleontologist George Olsen recovered the holotype (AMNH 6554), or name-bearing specimen, of Alectrosaurus, a nearly complete right hindlimb. This included the distal end of the right femur, the tibia, the fibula, the astragalus, the calcaneum, an incomplete right pes, three metatarsals of the left hind foot, two manual unguals, a manus, and the distal end of the pubis known as the pubic foot. On May 4th, Olsen discovered AMNH 6368 approximately 30 meters away from his first find. This specimen included a right humerus, two incomplete manual digits, four fragmentary caudal vertebrae, and other poorly preserved material. These discoveries were made at the Iren Dabasu Formation in what is now the Inner Mongolia Autonomous Region (Nei Mongol Zizhiqu) of the People's Republic of China. The age of this geologic formation is not clear, but is commonly cited as the Campanian stage of the Late Cretaceous Period, about 83 to 72 million years ago. More material, including comparable hind limb material as well as skull and shoulder elements, has been referred to Alectrosaurus. These fossils were found in the Bayan Shireh Formation of Outer Mongolia, a formation which is also of uncertain age. It may possibly extend into the early Campanian, but recent estimates suggest it was deposited from Cenomanian through Santonian times. Iren Dabasu and Bayan Shireh dinosaur faunas are similar, but van Itterbeecka et al. claimed that the Iren Dabasu is probably Campanian-Maastrichtian in age and possibly correlated with the Nemegt Formation, so it is not surprising that a species of Alectrosaurus would be found there. Furthermore, several more partial skeletons may have been found in both Inner and Outer Mongolia. Alectrosaurus was a medium sized, moderately built carnivorous dinosaur. The length of its tibia and femur are very close, in contrast to the majority of other tyrannosauroids, where the tibia is longer. The hind foot (and ankle) are also closer in size to the tibia than in most other tyrannosauroids, where the hind foot is usually longer. In 1933 Charles Gilmore examined the available material and concluded that AMNH 6554 and AMNH 6368 were syntypes belonging to the same genus. He based this on his observation that the manual unguals from both specimens were morphologically similar. Observing similarities with the hindlimbs of specimen AMNH 5664 Gorgosaurus sternbergi, he classified this new genus as a "Deinodont", a term that is now considered equivalent to tyrannosaurid. Due to its fragmentary nature, there is presently very little confidence in restoring its relationships with other tyrannosauroids and many recent cladistic analyses have omitted it altogether. One study recovered Alectrosaurus at no less than eight equally parsimonious positions in a tyrannosauroid cladogram. Alectrosaurus was originally characterized as a long-armed theropod, but Mader and Bradley (1989) observed that the forelimbs (AMNH 6368) did not belong to this individual and assigned them to the segnosauridae. The remaining material, AMNH 6554 represents the hind limb of a true tyrannosauroid, and were assigned as the lectotype for Alectrosaurus olseni. Mader and Bradley also described and assigned caudal vertebrae AMNH 21784 to this genus. These researchers concluded that Alectrosaurus was closely related to Maleevosaurus novojilov based on hind limb proportions. Some paleontologists have considered Alectrosaurus olseni to be a species of Albertosaurus. The Bayan Shireh material may or may not belong to this genus, and needs further study. One cladistic analysis showed that the two sets of specimens group together exclusive of any other taxa, so they are probably at least closely related, if not the same species. ( Read More ) Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| 01:02 pm [industrialterro] [Link] |
Albertosaurus (часть 1) Альбертоза́вр (Albertosaurus) — род теропод из семейства тираннозаврид. Существовал в западной части Северной Америки (от Аляски до Техаса) во время позднего мелового периода (76—65 миллионов лет назад). Ареал типового вида Albertosaurus sarcophagus был ограничен областью современной канадской провинции Альберты, отсюда и произошло его название. Как и все тираннозавриды, альбертозавр был двуногим хищником с крошечными двупалыми передними лапками и массивной головой с десятками больших острых зубов. Считается, что альбертозавр был доминирующим хищником в своей среде обитания. Будучи относительно крупным для теропода, по размерам он значительно уступал своему более известному родственнику тираннозавру — средний вес особей альбертозавра был менее двух тонн. Изучение вида ведётся с 1884 года, всего были обнаружены останки более тридцати особей, что позволило учёным изучить анатомию альбертозавра лучше других представителей семейства. Открытие двадцати особей различного возраста на одном участке (что может являться свидетельством стайного поведения) позволяет исследовать онтогенез и популяционную биологию альбертозавра, в отличие от большинства других динозавров. По размеру альбертозавр был меньше самых крупных тираннозаврид, например, тарбозавра и тираннозавра. Типичная взрослая особь достигала 9 метров в длину, некоторые старые особи могли превышать 10 метров. Вес взрослых альбертозавров, вычисленный на основе ископаемых останков разными методами, составлял от 1,2 до 1,7 тонны. Массивный череп альбертозавра, достигавший 1 метра в длину, поддерживала короткая S-образная шея. Широкие отверстия в черепе (окна) снижали вес головы, а также предоставляли пространство для челюстных мышц и сенсорных органов. Длинные челюсти альбертозавра содержали более 60 длинных, похожих на бананы зубов (более крупные тираннозавриды имели меньшее количество зубов). В отличие от большинства других теропод, альбертозавр и другие тираннозавриды были гетеродонтами, то есть имели разные виды зубов, в зависимости от их расположения во рту. Зубы, находившиеся на резцовой кости, были меньше остальных, располагались более тесно и имели D-образную форму в сечении. Над глазами имелись короткие костяные гребни, которые при жизни могли быть яркого цвета и использоваться при ухаживании для привлечения партнёра. Все тираннозавриды, в том числе альбертозавр, имеют сходное строение тела. Как и большинство тероподов, альбертозавр был бипедальным и уравновешивал тяжёлую голову и корпус очень длинным хвостом. Передние конечности верхнемеловых тираннозаврид были чрезвычайно мелкими и сохранили только два когтя. Задние конечности были длинными и заканчивались четырьмя пальцами с мощными когтями. Короткий первый палец задней конечности не участвовал в передвижении, только три других соприкасались с землёй, третий (средний) был длиннее остальных. Считается, что альбертозавр мог развивать скорость от 14 до 21 км/ч. Альбертозавр был представителем семейства тираннозаврид, подсемейства альбертозаврин. Его самый близкий «родственник» — обнаруженный в породах более позднего возраста горгозавр (иногда называемый Albertosaurus libratus). Эти два рода — единственные известные представители подсемейства альбертозаврин, но могли существовать и другие. Палеонтолог Томас Хольц в 2004 году предположил, что аппалачиозавр тоже относился к альбертозавринам, но в своей более поздней работе уже не рассматривал аппалачиозавра как представителя этого подсемейства, и эта точка зрения поддерживается и другими учёными. Другим подсемейством тираннозаврид являются тираннозаврины, к которым относятся дасплетозавр, тарбозавр и тираннозавр. По сравнению с тираннозавринами, альбертозаврины имели более стройное тело, пропорционально меньшие черепа и кости таза и более длинные большеберцовые и таранные кости, а также кости плюсны и пальцев. Название этому виду динозавров дал известный американский геолог, евгеник и палеонтолог Генри Осборн в 1905 году в одностраничном комментарии в конце своего описания Tyrannosaurus rex. Название «альбертозавр» происходит от названия канадской провинции Альберта, на территории которой впервые были найдены кости. Родовое название также включает в себя греческое слово σαῦρος (saurus) — ящер. Название типового вида «sarcophagus» означает «хищник», «плотоядный» и имеет то же происхождение, что и слово «саркофаг» (от древнегреческого σάρξ (sarx) — плоть и φαγεῖν (phagein) — поедать). Науке известно более тридцати скелетов этого динозавра всех возрастов. Типовым экземпляром является фрагментированный череп, обнаруженный в 1884 году в отложениях карьера Хорсшу одноимённого каньона, расположенного рядом с рекой Ред-Дир на территории современной провинции Альберта. Этот экземпляр и меньший череп, вместе с некоторым скелетным материалом, были обнаружены геологической экспедицией во главе с известным геологом Джозефом Тирреллом. Эти два черепа были отнесены к ранее описанному виду Laelaps incrassatus Эдвардом Коупом в 1892 году, несмотря на то, что название Laelaps было ранее дано роду клещей и было изменено на Dryptosaurus в 1877 году Чарлзом Маршем. Коуп отказался признать новое название, созданное его главным научным противником; несмотря на это, Лоуренс Ламбе в 1904 году описал эти черепа, отнеся их к виду Dryptosaurus incrassatus, а не Laelaps incrassatus. Чуть позже Генри Осборн указал на то, что описание Dryptosaurus incrassatus было основано на нескольких основных зубах тираннозаврид, поэтому два черепа из Альберты не могут с уверенностью быть отнесены к этому виду. Эти черепа также значительно отличались от останков типового вида дриптозавра — Dryptosaurus aquilunguis. Таким образом, Осборн в 1905 году выделил на основании этих черепов новый вид — Albertosaurus sarcophagus. При классификации он полагался на описание, созданное Ламбе на год раньше. Оба экземпляра (КМП 5600 и КМП 5601) были выставлены в Канадском музее природы в Оттаве. В 1910 году американский палеонтолог Барнум Браун обнаружил множество костей, принадлежавших большой группе альбертозавров в другом карьере, рядом с рекой Ред-Дир. Из-за большого количества образцов и недостатка времени экспедиция Брауна не откопала все экземпляры, но всё же ей удалось собрать из костеносного слоя все особи, которые можно было идентифицировать на тот момент. В тот год было обнаружено семь разных наборов костей стопы, а также два образца, несоответствующих другим по размерам и, возможно, принадлежащих детёнышам. Все они находятся в коллекции Американского музея естественной истории. Королевский Тиррелловский палеонтологический музей возобновил экспедицию в этот район (Сухой остров Буффало-Джамп) в 1997 году. Раскопки проводились с 1997 по 2005 год. В результате были обнаружены кости ещё 13 особей, среди них двухлетнего детёныша и очень старого альбертозавра, длина которого была оценена в 10 метров. Ни один скелет из найденных экземпляров не был полным. Кости были выставлены в Тирелловском музее и Американском музее естественной истории. В 1913 году палеонтолог Чарльз Штернберг обнаружил скелет тираннозаврида в формации Парк динозавров в Альберте. В следующем 1914 году Лоуренс Ламбе дал этому динозавру название Gorgosaurus libratus. Позже в Альберте и американском штате Монтана были найдены другие останки горгозавров. Обнаружив, что горгозавр и альбертозавр имеют слишком мало отличий, Дейл Рассел предложил считать горгозавра младшим синонимом альбертозавра, а так как альбертозавр был открыт и назван первым, в 1970 году горгозавр был переименован в Albertosaurus libratus, из-за чего временной диапазон обитания альбертозавра увеличился на несколько миллионов лет, а ареал простирался на сотни километров южнее, чем это считалось раньше. В 2003 году палеонтолог Фил Карри изучил черепа нескольких тираннозаврид, в том числе и горгозавра и альбертозавра, и выяснил, что они имеют всё же несколько больше различий, чем это считалось ранее. На сегодняшний день вопрос о том, использовать один или два рода, не решён окончательно, так как горгозавр и альбертозавр являются очень близкими таксонами. Признавая это, Карри, однако, рекомендовал, чтобы альбертозавр и горгозавр считались отдельными родами, поскольку отличий между ними не меньше, чем между дасплетозавром и тираннозавром, которых почти всегда можно отличить друг от друга. Кроме того, недавно были обнаружены новые скелеты альбертозаврин в штате Нью-Мексико и на Аляске, и Карри предположил, что вопрос о альбертозавре-горгозавре может быть полностью разъяснён, как только они будут изучены. Большинство исследователей поддерживают точку зрения Карри, но есть и противники этой гипотезы. В 1928 году Уильям Паркс описал новый вид — Albertosaurus arctunguis на основании частичного скелета, раскопанного у реки Ред-Дир, но с 1970 года этот вид считается идентичным Albertosaurus sarcophagus. Этот экземпляр (КМО 807) хранится в Королевском музее Онтарио в Торонто. С тех пор в Альберте было обнаружено ещё шесть экземпляров, причисленных к этому виду. Ископаемые, причисленные к Albertosaurus sarcophagus, были также обнаружены в американских штатах Монтана, Нью-Мексико и Вайоминг, но они, вероятно, не относятся к этому виду, а возможно, и к роду альбертозавр. Albertosaurus megagracilis был описан на основании небольшого скелета, найденного в геологической формации Хелл-Крик. В 1995 году он был выделен в отдельный род — Dinotyrannus, который позже стали описывать как принадлежащий подростковой особи тираннозавра рекса. Репродукции (1, 2, 3, 4, 5, 6):
Размеры тела в сравнении с человеком: Ископаемые останки и реплики (1, 2, 3, 4, 5, 6):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| 12:50 pm [industrialterro] [Link] |
Albertosaurus (часть 2) До нашего времени сохранились останки почти всех возрастных групп альбертозавра. С помощью гистологического исследования можно определить возраст животного в момент его смерти. Сравнив останки особей одного вида, но разного возраста, можно определить среднюю скорость роста этих животных. Самый молодой из известных альбертозавров — двухлетний, он был обнаружен в Буффало-Джамп, весил приблизительно 50 килограммов, а в длину достигал чуть более двух метров. Десятиметровый экземпляр, обнаруженный в том же карьере, является самым старым из известных — он умер в возрасте двадцати восьми лет. Наибольший темп роста альбертозавра приходился на период от четырёх до шестнадцати лет, как и у некоторых других тираннозаврид. В этот период жизни альбертозавры набирали 122 килограмма ежегодно, пока не достигали 1,3 тонны. Впрочем, согласно другим исследованиям, вес взрослых альбертозавров был больше, и темпы роста тоже могли быть больше. У тираннозаврид, близких по размерам к альбертозавру, темпы роста были схожи, а более крупные, такие как тираннозавр, наоборот, росли почти в пять раз быстрее (601 килограмм ежегодно). Конец фазы быстрого роста у альбертозавров предполагал достижение половой зрелости; хотя они и продолжали расти до конца жизни, но гораздо медленнее. Раннее половое созревание в процессе роста типично для мелких и крупных динозавров, как и для большинства млекопитающих, таких как люди и слоны. С другой стороны, ближайшие родственники динозавров, птицы, достигают половой зрелости лишь по завершении процесса роста. Чаще всего встречаются останки особей альбертозавров в возрасте четырнадцати лет и старше. Более молодых животных находят редко по нескольким причинам — в основном из-за того что кости молодых животных сохраняются хуже, чем кости взрослых, а также из-за того что искателям окаменелостей их сложнее заметить. Несмотря на то что молодые альбертозавры были достаточно крупными животными, их останков обнаружено гораздо меньше, чем взрослых. Предполагается, что такая ситуация сложилась вследствие хороших условий жизни для детёнышей, из-за чего они умирали реже взрослых. Согласно общей теории, уже по достижении всего двух лет детёныши альбертозавра были крупнейшими хищниками своего ареала, они также были гораздо быстрее, чем все их жертвы. Из-за этого даже у молодых альбертозавров не было врагов. Это и привело к уменьшению смертности детёнышей и, соответственно, к уменьшению количества их останков. Останки двенадцатилетних альбертозавров встречаются в два раза чаще, чем более молодых, а достигших половой зрелости четырнадцатилетних и старше — ещё в два раза больше. Возможно, по достижении половой зрелости усиливалась внутривидовая борьба за самку или территорию. Более высокая смертность взрослых объясняет большее количество сохранившихся останков. Очень крупные особи также встречаются очень редко, поскольку лишь немногим удавалось прожить столь долго, чтобы достигнуть таких размеров. Схема, в которой высокий уровень смертности наблюдается среди детёнышей, низкий среди подростков, повышенный среди взрослых особей и крайне небольшое количество животных способно дожить до глубокой старости, свойственна большинству современных крупных животных, таких как носороги, африканский буйвол или слон, а также некоторым тираннозавридам, за исключением, разумеется, альбертозавра, детёныши которого умирали редко. В связи с пробелами в палеонтологической летописи, выводы об уровне смертности не могут быть более точными, в том числе и потому что более двух третей всех останков альбертозавров были найдены в одном месте. В Буффало-Джамп были обнаружены останки двадцати двух альбертозавров, что является самым крупным массовым захоронением тероподов после массового захоронения аллозавров в карьере динозавров Кливленд-Ллойд в Юте. В захоронении присутствуют останки одного старого взрослого, восьми взрослых между семнадцатью и двадцатью тремя годами, семи подростков в возрасте от двенадцати до шестнадцати лет и шести детёнышей между двумя и одиннадцатью годами, которые ещё не достигли фазы быстрого роста. Почти полное отсутствие останков травоядных животных и одинаковый уровень сохранности останков альбертозавров в карьере навели Фила Карри на мысль, что это захоронение не было ловушкой для хищников, как смоляные ямы на ранчо Ла-Брея в Калифорнии, и что все сохранившиеся животные умерли в одно время. Карри считает, что это свидетельство стайного поведения. Другие учёные более скептичны, отмечая, что, возможно, животных согнала вместе засуха или их тела были снесены в одно место наводнением. Есть многочисленные свидетельства стадного образа жизни среди растительноядных динозавров, в том числе цератопсов и гадрозавров. Однако массовые захоронения хищных динозавров редки. Маленькие тероподы, такие как дейноних, целофиз и мегапнозавр (синтарс), были найдены в больших скоплениях, как и крупные хищники аллозавр и мапузавр. Есть также доказательства стайного поведения среди других тираннозаврид. Фрагментарные остатки детёнышей были найдены рядом с тираннозавром по прозвищу «Сью», хранящимся в музее естественной истории имени Филда в Чикаго, а в костеносном слое геологической формации Ту-Медисин в Канаде были найдены останки трёх дасплетозавров рядом с несколькими гадрозаврами. Эти находки могут подтвердить, что альбертозавр также вёл социальный образ жизни, хотя некоторые вышеупомянутые скопления могут быть временными или неестественными. Но этому может быть и другое объяснение. По крайней мере, некоторые из этих находок представляют собой проявления массового каннибализма, как у комодского варана. Карри также считает, что доказательством стайного образа жизни альбертозавров может быть их способ охоты. Пропорции ног молодых альбертозавров были сопоставимы с таковыми у орнитомимид, которые, пожалуй, были самыми быстрыми среди динозавров. Они, по всей видимости, были быстрее любых травоядных своего времени. Карри выдвинул гипотезу, что младшие члены стаи отбивали жертву от стада и загоняли её к взрослым особям, которые были сильнее детёнышей, но медлительней. Животные подросткового возраста, предположительно, вели образ жизни, отличавшийся от поведения своих взрослых родственников, заполняя экологическую нишу между ними и мелкими тероподами, крупнейшие из которых весили вдвое меньше взрослого альбертозавра. Подобная ситуация наблюдается у современных комодских варанов, которые начинают жизнь как маленькие насекомоядные животные, а потом становятся доминирующими хищниками. Тем не менее, полностью воссоздать поведение вымерших животных на основании окаменелостей невозможно. Все идентифицируемые окаменелости Albertosaurus sarcophagus были обнаружены в формации каньона Хорсшу. Эта геологическая формация относится к маастрихтскому ярусу мелового периода, возрастом 70—73 миллиона лет. Непосредственно ниже этой формации находятся сланцы морской формации Бирпо, которая является частью более обширной морской формации Сиуэй. В конце мелового периода море отступало, климат становился холоднее, уровень моря уменьшался, обнажая землю, которая раньше находилась под водой. Этот процесс, однако, не был гладким: из-за сильных бурь на море часть суши в районе каньона Хорсшу частично затоплялась, но в конце концов море всё-таки отступило. Из-за постоянно изменяющегося уровня моря в формации Хорсшу было много прибрежных лагун, болот и илистых осушек. Многочисленные угольные пласты представляют древние торфяные болота. Как и в случае большинства других позвоночных в этой формации, останки альбертозавра были найдены в отложениях доисторических дельт и пойм больших рек. Доисторическая фауна формации Хорсшу хорошо известна, поскольку окаменелости позвоночных, включая динозавров, присутствуют здесь в массовых количествах. Фауну рыб составляли акулы, скаты, осетры, ильные рыбы, панцирникообразные и подобные панцирниковым аспидохиноморфы. Фауна млекопитающих включала в себя многобугорчатых и сумчатых, таких как дидельфодон. В морских отложениях был обнаружен плезиозавр леуроспондил, а в отложениях пресноводной среды были найдены черепахи, хампсозавры и крокодилы, такие как лейдиозух и стангерохампс. Доминирующими представителями фауны были динозавры, особенно распространены были гадрозавры, которые составляют половину всех найденных здесь динозавров, к ним относились эдмонтозавр, зауролоф и гипакрозавр. Цератопсы и орнитомимиды также были очень распространены, составляя треть найденных здесь динозавров. Наряду с намного более редко встречающимися анкилозаврами и пахицефалозаврами, эти растительноядные динозавры были добычей для множества плотоядных тероподов, таких как троодонтиды, дромеозавриды и кенагнатиды. Взрослые альбертозавры были доминирующими хищниками в этой среде и находились на вершине пищевой пирамиды, а промежуточные ниши между мелкими тероподами и взрослыми альбертозаврами, возможно, занимали юные альбертозавры. William Abler observed in 2001 that Albertosaurus tooth serrations resemble a crack in the tooth ending in a round void called an ampulla. Tyrannosaurid teeth were used as holdfasts for pulling meat off a body, so when a tyrannosaur pulled back on a piece of meat, the tension could cause a purely crack-like serration to spread through the tooth. However, the presence of the ampulla distributed these forces over a larger surface area, and lessened the risk of damage to the tooth under strain. The presence of incisions ending in voids has parallels in human engineering. Guitar makers use incisions ending in voids to, as Abler describes, "impart alternating regions of flexibility and rigidity" to the wood they work with. The use of a drill to create an "ampulla" of sorts and prevent the propagation of cracks through material is also used to protect aircraft surfaces Abler demonstrated that a plexiglass bar with incisions called "kerfs" and drilled holes was more than 25% stronger than one with only regularly placed incisions. Unlike tyrannosaurs, ancient predators like phytosaurs and Dimetrodon had no adaptations to prevent the crack-like serrations of their teeth from spreading when subjected to the forces of feeding. In 2009, researchers hypothesized that smooth-edged holes found in the fossil jaws of tyrannosaurid dinosaurs such as Albertosaurus were caused by a parasite similar to Trichomonas gallinae, which infects birds. They suggested that tyrannosaurids transmitted the infection by biting each other, and that the infection impaired their ability to eat food. In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. They found that only one of the 319 Albertosaurus foot bones checked for stress fractures actually had them and none of the four hand bones did. The scientists found that stress fractures were "significantly" less common in Albertosaurus than in the carnosaur Allosaurus. ROM 807, the holotype of A. arctunguis (now referred to A. sarcophagus), had a 2.5 by 3.5 cm deep hole in the iliac blade, although the describer of the species did not recognize this as pathological. The specimen also contains some exostosis on the fourth left metatarsal. In 1970, two of the five Albertosaurus sarcophagus specimens with humeri were reported by Dale Russel as having pathological damage to them. In 2010, the health of the Dry Island Albertosaurus assembly was reported upon. Most specimens showed no sign of disease. On three phalanges of the foot strange bony spurs, consisting of abnormal ossifications of the tendons, so-called enthesophytes, were present, their cause unknown. Two ribs and a belly-rib showed signs of breaking and healing. One adult specimen had a left lower jaw showing a puncture wound and both healed and unhealed bite marks. The low number of abnormalities compares favourably with the health condition of a Majungasaurus population of which it in 2007 was established that 19% of individuals showed bone pathologies. Тираннозаври́ды (Tyrannosauridae) — семейство тероподовых динозавров, вымерших в конце мелового периода (65 миллионов лет назад). Обитали в Азии и в Северной Америке. Известны с раннего мела. Подразделяются на два подсемейства — длинномордых Albertosaurinae и короткомордых Tyrannosaurinae. К тираннозавридам относят такие вымершие роды, как тарбозавр (длина 10 метров), дасплетозавр (длина 8,5 метров), монгольский алиорам (длина 6 метров), алтайский алиорам и другие. Самым известным представителем надсемейства является тираннозавр (занимает 4 место по размеру среди тероподов). Тираннозавр имел длину до 14 метров, в высоту достигал 5 метров, а весил около 7 тонн. Тираннозавриды главным образом характеризуются общими скелетными структурами, особенно характерны структуры бедренных и соединение носовых костей. Массивные головы и миниатюрные передние лапы с двумя пальцами делают этих меловых двуногих хищников легко узнаваемыми. Ранние тираннозавриды не имели характерного широкого, массивного черепа и редуцированных передних конечностей, как их потомки. Эти ранние виды обычно имели маленькие или средние черепа относительно размера их тела, были легче, стройнее, имели длинные, трёхпалые передние конечности. У них не было загнутых (бананоподобных) зубов, типичных для тираннозавровых. Внешне они наверняка были больше похожи на целурозавров чем на поздних тираннозавров. Некоторые исследователи предлагают, что некоторые или все известные «целурозавры» могут быть ранними тираннозавридами, хотя детальное изучение ещё не опровергло и не подтвердило эту теорию. Тираннозавриды — главным образом обитатели Евразии, а в Северную Америку они проникли лишь в конце мела, когда образовался сухопутный мост через Берингов пролив — Берингия. Об этом свидетельствуют находки костей в префектуре Исикава, Япония, которые предположительно принадлежат данному семейству и датированы возрастом около 140 млн лет. В Северной Америке тираннозавриды были найдены на территории Канады и США. В 2011 году был описан новый род Teratophoneus, найденный в штате Юта в слоях датируемых поздним мелом — 75 млн лет. Репродукции (7, 8, 9, 10, 11, 12): Размеры тела в сравнении с человеком: Ископаемые останки и реплики (7, 8, 9, 10, 11, 12): Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| April 18th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 08:06 pm [industrialterro] [Link] |
Albertonykus Albertonykus borealis — вид динозавров-теропод из семейства Alvarezsauridae, живших во время позднего мелового периода в районе нынешней Северной Америки. Вид был описан в 2008 году Филом Карри и Ником Лонгричем. Первая часть названия рода относится к канадской провинции Альберта, вторая часть «nykus» — искажённое греческое «νυχος» — коготь. Видовой эпитет означает — «северный». Окаменелости датируются началом маастрихта, 70 миллионов лет назад. Голотип TMP 2001.45.91 состоит из левой голени. Есть и другие фрагменты костей, которые принадлежат по меньшей мере двум особям. На месте также были найдены кости большого теропода — альбертозавра. Albertonykus был теплокровным пернатым динозавром, длиной чуть менее одного метра. По данным кладистического анализа, Albertonykus является сестринским таксоном клады Mononykinae. В статье с описанием динозавра данная особенность рассматривается как подтверждение гипотезы, что альваресзавриды возникли в Южной Америке, а затем через Северную Америку и сухопутный мост на территории современного Берингова пролива проникли в Азию. Albertonykus является самым ранним среди известных динозавров семейства Alvarezsauridae Северной Америки. Albertonykus (meaning "Alberta claw") is a genus of alvarezsaurid dinosaur from Maastrichtian-age (Upper Cretaceous) rocks of the Horseshoe Canyon Formation of Alberta, Canada. It is known from forelimb and hindlimb remains from multiple individuals. All but two of the specimens come from a bonebed dominated by Albertosaurus. This bonebed is located at the top of Unit 4 of the Horseshoe Canyon Formation, which dates to about 68.5 million years ago. Albertonykus is interpreted as having fed on wood-nesting termites because the forelimbs appear to be specialized for digging, but are too short for burrowing. Albertonykus is the earliest-known North American alvarezsaurid; isolated remains of alvarezsaurids are known from later rock units in Montana and Wyoming (USA). The type species is A. borealis, described by Nicholas Longrich and Philip Currie in a paper published in 2009 (which was available online as a preprint in 2008). The specific name (borealis) means "north". Until the 1900s it was unknown that dinosaurs like Albertonykus existed. In 1910, paleontologist Barnum Brown and his crew found a site filled with the bones of the tyrannosaur Albertosaurus sarcophagus. They unearthed the remains of at least nine different specimens of Albertosaurus in the single site, but stopped digging after just two weeks. The quarrying again produced the remains of over 20 specimens of Albertosaurus, and the quarry has yet to be fully excavated. It is not known why so many tyrannosaurs were found in one place: paleontologist Nick Longrich speculated that the site was part of a "predator trap", where food drew predators to their deaths. Close to a dozen arms and leg bones belonging to an unknown species of Albertosaurus were recovered in Alberta, Canada in 2002 by a team of scientist led by Philip J. Currie of the University of Alberta. The bones were then stored at the Royal Tyrrell Museum in Alberta. In 2005 Nick Longrich, a paleontologist at the University of Calgary in Canada found the fossils while comparing Albertosaurus claws to other dinosaur species. Longrich then analyzed the fossils found in the Albertosaurus bonebed in Dry Island Provincial Park to find that they dated back 70 million years and represented a completely unknown species. Although fragments of Albertonykus had been discovered before, they had never been identified as a separate species. A chain of new discoveries found in Asia and South America exposed the existence of these previously unknown theropod dinosaurs. Albertonykus is the smallest known alvarezsaurid ever discovered in North America, measuring only 70 centimeters (2.5 ft) in length. Alvarezsaurs typically had slender hind legs, long rigid tails, and unusually short arms that were powerfully built to support a large claw. Although no skull bones of Albertonykus have been found, related animals from Mongolia show that they likely had long, slender snouts filled with tiny teeth similar to those of armadillos and anteaters. It is likely that Albertonykus ate insects, using its large thumb claw to tear open rotten logs to find its prey. As in other alvarezsaurs, the forelimbs of Albertonykus were specialized for digging, but were too short to permit burrowing. Unfortunately at this time the skeleton of Albertonykus is not complete, but its Mongolian relatives give us a distinct idea of what the rest of the skeleton would have looked like. Ants were not an important part of the ecosystem during the Cretaceous, and mound-building termites do not appear until the Eocene. This leaves the possibility that Albertonykus preyed on wood-nesting termites. This hypothesis was tested by examining petrified wood from the Horseshoe Canyon Formation, where Albertonykus was found. The wood found there frequently contains borings, which resemble those of termites. Albertonykus is a member of an unusual theropod group known as the Alvarezsauridae and is one of only a few alvarezsaurid fossils that has been found outside of South America and Asia. After a phylogenetic analysis, it was shown that Albertonykus is the sister taxon of the Asian clade Mononykinae, which supported the hypothesis that the alvarezsaurs originated in South America, and then dispersed to Asia through North America. The unearthing of Albertonykus provided important information into the biology of the Alvarezsauridae. Tags: Вымершие рептилии, Мел, авеметатарзалии, альваресзавриды, альваресзавры, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, манирапторы, тероподы, целурозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| April 15th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 08:56 pm [industrialterro] [Link] |
Albertaceratops Альбертацератопс (Albertaceratops; букв. «рогатая морда из Альберты») — род птицетазовых динозавров из семейства цератопсид, живших в раннем меловом периоде (около 83,5—70,6 миллионов лет назад) на территории нынешней Северной Америки. Окаменелости динозавра были найдены на территории провинции Альберта (Канада) и штата Монтана (США). Впервые описан палеонтологом Ryan в 2007 году. Всего известно два скелета Albertaceratops. Представлен одним видом — Albertaceratops nesmoi. Albertaceratops (meaning "Alberta horned face") was a genus of centrosaurine horned dinosaur from the middle Campanian-age Upper Cretaceous Oldman Formation of Alberta, Canada. Albertaceratops is known from a single complete skull (TMP.2001.26.1) found in August 2001 and skull and postcranial fragments. This genus is unusual in combining long brow horns with an otherwise centrosaurine skull, as centrosaurines normally possess short brow horns. Over its nose was a bony ridge, and on its frill were two large outwardly-projecting hooks. A phylogenetic analysis carried out by its describer, Michael J. Ryan, found it to be the most basal centrosaurine. Additional specimens were reported from a bonebed in the Judith River Formation of Montana, which is equivalent to the Oldman Formation and differentiated only by the United States/Canadian border. However, further study showed these remains to come from a chasmosaurine, Medusaceratops. Both ceratopsids lived during the same time period, about 77.5 million years ago. The specific name, A. nesmoi, is derived from the name of Cecil Nesmo, a rancher living in Manyberries, Alberta, a town of less than 100 people located 71 km south of Medicine Hat. The rancher was thus honored in recognition of his efforts to aid fossil hunters. Репродукции (1, 2, 3, 4, 5, 6): Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморвф, динозавры, маргиноцефалии, неорнитискии, птицетазовые, центрозаврины, цераподы, цератопсиды, цератопсы | ||||||||||||||||||||||||||||||||||||||||
| 08:13 pm [industrialterro] [Link] |
Albalophosaurus Альбалофозавр (Albalophosaurus) — род птицетазовых динозавров подотряда цераподов, живших в раннем меловом периоде (около 145—140 миллионов лет назад), на территории нынешней Азии. Окаменелости теропода были найдены в геологической формации Kuwajima Formation в Японии. Впервые описан палеонтологами Ohashi и Barrett в 2009 году. Представлен одним видом — A. yamaguchiorum. В результате кладистического анализа было установлено, что Albalophosaurus относится к группе Cerapoda. Другие характеристики, такие как морфология зубов Albalophosaurus предполагают отнесение данного рода к Ornithopoda. Учитывая много недостающей информации исследователи ограничились размещением данного таксона Cerapoda указав его как incertae sedis. Albalophosaurus (meaning 'white crest lizard') is a genus of ceratopsian ornithischian dinosaur. It was described in 2009 from remains found in 1997 by Yoshinori Kobayashi from the Kuwajima Formation of central Japan, outcropping in Hakusan in the Ishikawa Prefecture. The holotype, SBEI 176, consists of cranial bones from an incomplete, disarticulated skull and left lower jaw thought to belong to a single individual. The type species is named A. yamaguchiorum. The generic name is derived from Latin albus, "white", and Greek λόϕος (lophos), "crest", a reference to the snow-covered crest of Mount Hakusan. The specific name honours Ichio Yamaguchi en Mikiko Yamaguchi, who discovered and prepared many fossils from the site. The exact age of the strata from which the remains of Albalophosaurus have been found is not known because of the lack of marine beds containing index fossils, but the Kuwajima Formation is known to have formed during the Early Cretaceous, most likely after the Berriasian and before the Barremian based on the ages of underlying and overlying formations. More recent studies suggest that the age of the Kuwajima Formation is most likely Valanginian—Hauterivian, although the exact age is still uncertain. Although Albalophosaurus was classified as a basal ceratopsian in a phylogenetic analysis conducted along with the description of the genus, only one ambiguous synapomorphy of the clade is present in the holotype, and none of the unambiguous synapomorphies that define Ceratopsia are present. Other characteristics, such as those of the dental morphology of Albalophosaurus, seem to suggest that the genus shares a relation to Ornithopoda. Thus the authors of the original description of the genus refer it to Cerapoda incertae sedis, and do not consider it to be a ceratopsian. Han et al., 2012 found Albalophosaurus to be a ceratopsian more derived than Micropachycephalosaurus, Yinlong, Stenopelix, and Chaoyangosaurus, but basal to a clade composed of Psittacosaurus and more derived ceratopsians. Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, птицетазовые, цераподы, цератопсы | ||||||||||||||||||||||||||||||||||||||||
| April 6th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 08:22 pm [industrialterro] [Link] |
Alaskacephale Alaskacephale was a genus of pachycephalosaurid dinosaur that lived in the late Campanian to Maastrichtian stages of the Late Cretaceous (around 80 to 69 million years ago). Alaskacephale was named by Robert Sullivan in 2006. The genus name refers to Alaska, where the fossil was found in the Prince Creek Formation. The species name, gangloffi, honors paleontologist Roland Gangloff. The only known specimen of A. gangloffi is the holotype, a nearly complete left squamosal with a characteristic array of polygonal nodes. The dimensions of this bone suggest that A. gangloffi was about half the size of Pachycephalosaurus wyomingensis or three quarters the size of Prenocephale prenes, and about the same size as Prenocephale edmontonensis and Prenocephale brevis. The specimen was previously described by Gangloff et al. (2005) as an unnamed pachycephalosaurid, possibly a Pachycephalosaurus. Gangloff et al. described the squamosal as having a suture with the quadrate, a feature previously described only in Pachycephalosaurus. Sullivan (2006) opined that this "suture" is instead a breakage point in both Alaskacephale and Pachycephalosaurus, so it could not be used to unite the two taxa. Аляска известна не только суровым климатом и потрясающей природой. Здесь располагается богатая формация Принс-Крик, которая, на радость палеонтологов всего мира, регулярно поставляет новые таксоны динозавров. Рассмотрим первого и пока единственного обнаруженного там пахицефалозавра. Латинское название Alaskacephale происходит от древнегреческого слова "голова" и "Аляска". Существовали аляскацефалы в конце мелового периода, около 72 – 68 миллионов лет назад (ранний маастрихтский ярус). Были распространены на территории Соединённых Штатов Америки, в Аляске. Ныне известен единственный вид – Alaskacephale gangloffi, соответственно являющийся типовым. Единственный обнаруженный экземпляр аляскацефала (соответственно, голотип) получил идентификационный номер UAM AK-493-V-001. Он представляет собой чешуйчатую кость с характерными рядами конусовидных выростов – части обрамления купола. Аналогичное имелось у североамериканского родича – пахицефалозавра (Pachycephalosaurus). Длина тела аляскацефала достигала 2,25 метров. Высота доходила до 0,9 метров, а весил этот ящер около 40 кг. Североамериканская свита Принс-Крик, с лихвой насыщенная водоёмами, в начале маастрихта стала прибежищем многих растительноядных динозавров от мала до велика. По соседству с небольшим аляскацефалом могли проживать даже внушительные эдмонтозавры (Edmontosaurus) и пахиринозавры (Pachyrhinosaurus). Кто же мог представлять для аляскацефала угрозу? Этот ящер наверняка обладал неплохой скоростью передвижения. В этой формации обнаружены троодоны (Troodon), заврорнитолесты (Saurornitholestes) и ещё один неидентифицированный дромеозаврид ("Dromaeosaurus albertensis"). Эти хищники были хорошо вооружены и достаточно быстры для охоты на аляскоцефала, тем более что они могли атаковать добычу группами. Пахицефалоза́вры (Pachycephalosauria) — инфраотряд травоядных динозавров из группы Marginocephalia. Название впервые было использовано Марианской и Осмольской в 1974 году, но до сих пор точного таксономического определения данной группе не дано. В 1998 году Пол Серено предложил следующее определение: все виды, которые менее тесно связаны с трицератопсом, чем с Pachycephalosaurus. Вагнер в 2004 году в своём обзоре The Dinosauria дал другое определение: все виды, связанные ближе с Pachycephalosaurus wyomingensis чем с Triceratops horridus. В 2005 году Серено дал более сложное определение: все виды, более тесно связанные с Pachycephalosaurus wyomingensis, чем с Heterodontosaurus foxii, Ankylosaurus magniventris, Stegosaurus armatus, Iguanodon bernissartensis и Ceratops montanus. Выделение данной группы осложняется из-за многих неопределённостей внутренних взаимоотношений видов в отряде птицетазовых динозавров. Представители группы обитали с берриасского по маастрихтский века (146 до 65 млн. лет назад) на территории Азии и Северной Америки. Другие известные виды — Stygimoloch и Dracorex, которые иногда рассматриваются в качестве синонима рода Pachycephalosaurus. ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, пахицефалозавры, птицетазовые, цераподы | ||||||||||||||||||||||||||||||||||||||||
| March 24th, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 04:37 pm [industrialterro] [Link] |
Alamosaurus Аламозавр (Alamosaurus) — род ящеротазовых динозавров инфраотряда завропод. Включает один вид — Alamosaurus sanjuanensis. Эти ящеры жили в верхнем мелу (около 70—65 млн лет назад) на территории Северной Америки. Они достигали длины 21 м и веса 30 тонн. У аламозавра, как и у других завропод, была длинная шея и не менее длинный хвост, заканчивающийся кнутоподобным сужением. Вопреки популярному утверждению, этот динозавр не был назван в честь миссии Аламо в Техасе или произошедшей там в 1836 году битвы. Голотип этого вида был обнаружен в Нью-Мексико, и только после этого другие экземпляры были найдены и в Техасе. Родовое название дано по имени геологической свиты Ojo Alamo, в породах которой нашли первый экземпляр, а видовое — по имени округа Сан-Хуан штата Нью-Мексико, где была сделана эта находка. Аламозавра уверенно относят к группе титанозавров, в пределах которой он считается продвинутым родом. Но его родственные связи в пределах этой группы остаются неясными. Некоторые авторы включают его вместе с родом Opisthocoelicaudia в подсемейство Opisthocoelicaudinae семейства Saltasauridae. Согласно другим исследованиям, аламозавр — сестринская группа рода Pellegrinisaurus, и в состав Saltasauridae они не входят. Некоторые учёные отмечают сходство аламозавра с Neuquensaurus и Trigonosaurus. Alamosaurus (meaning "Ojo Alamo lizard") is a genus of titanosaurian sauropod dinosaurs, containing a single species, Alamosaurus sanjuanensis, from the late Cretaceous Period of what is now southern North America. It was a large quadrupedal herbivore. Isolated vertebrae and limb bones indicate that it reached sizes comparable to Argentinosaurus and Puertasaurus, which would make it the largest dinosaur known from North America. Its fossils have been recovered from a variety of geological formations spanning the Maastrichtian stage (late Edmontonian-Lancian land vertebrate ages) of the Late Cretaceous. Specimens of a juvenile Alamosaurus sanjuanensis have been recovered from only a few meters below the Cretaceous-Paleogene boundary in Texas, making it among the last surviving non-avian dinosaur species. The vertebrae from the middle part of its tail had elongated centra. Alamosaurus had vertebral lateral fossae that resembled shallow depressions. Fossae that similarly resemble shallow depressions are known from Saltasaurus, Malawisaurus, Aeolosaurus, and Gondwanatitan. Venenosaurus also had depression-like fossae, but its "depressions" penetrated deeper into the vertebrae, were divided into two chambers, and extend farther into the vertebral columns. Alamosaurus had more robust radii than Venenosaurus. Long thought to have been unarmored, recent discoveries indicate that Alamosaurus was armored like other lithostrotians, including Saltasaurus. Alamosaurus remains have been discovered throughout the southwestern United States. The holotype was discovered in the Naashoibito Member of the Ojo Alamo Formation (or Kirtland Formation under a different definition) of New Mexico which was deposited during the Maastrichtian stage of the Late Cretaceous Period. Bones have also been recovered from other Maastrichtian formations, like the North Horn Formation of Utah and the Black Peaks, El Picacho and Javelina Formations of Texas. Gilmore originally described a scapula (shoulder bone) and ischium (pelvic bone) in 1922. In 1946, he found a more complete specimen in Utah, consisting of a complete tail, a right forelimb complete except for the tips of the toes, and both ischia. Since then, many other bits and pieces from Texas, New Mexico, and Utah have been referred to Alamosaurus, often without much description. The most completely known specimen is a recently discovered juvenile skeleton from Texas, which allowed educated estimates of length and mass. One specimen of Alamosaurus, "USNM 15660", was first uncovered in 1937. Although originally found by Gilmore, it was first recognized by Michael Brett-Surman. It has been catalogued under the same number as a more complete and well-known specimen of Alamosaurus, which was found in very close proximity based on bone impressions. In 2015 it was realized that the specimen, which had been previously described, also included osteoderms, the first confirmation they existed on Alamosaurus. No skull material is known, except for a few slender teeth. Contrary to popular assertions, this dinosaur is not named after the Alamo in San Antonio, Texas, or the battle that was fought there. The holotype, or original specimen, was discovered in New Mexico and, at the time of its naming, Alamosaurus had not yet been found in Texas. Instead, the name Alamosaurus comes from Ojo Alamo, the geologic formation in which it was found and which was, in turn, named after the nearby Ojo Alamo trading post (since this time there has been some debate as to whether to reclassify the Alamosaurus-bearing rocks as belonging to the Kirtland Formation or whether they should remain in the Ojo Alamo Formation). The term alamo itself is a Spanish word meaning "poplar" and is used for the local subspecies of cottonwood tree. The term saurus is derived from saura (σαυρα), Greek for "lizard" and is the most common suffix used in dinosaur names. There is one species (A. sanjuanensis), which is named after San Juan County, New Mexico, where the first remains were found. Both genus and species were named by Smithsonian paleontologist Charles W. Gilmore in 1922. Alamosaurus fossils are most notably found in the Naashoibito member of the Ojo Alamo Formation (dated to between about 69-68 million years old) and in the Javelina Formation, though the exact age range of the later has been difficult to determine. A juvenile specimen of Alamosaurus has been reported to come from the Black Peaks Formation, which overlies the Javelina in Big Bend, Texas, and which straddles the Cretaceous-Paleogene boundary. The Alamosaurus specimen was reported to come from a few meters below the boundary, dated to 66 Ma ago, though the position of the boundary in this region is uncertain. Only one geological site in the Javelina Formation has thus far yielded the correct rock types for radiometric dating. The outcrop, situated in the middle strata of the formation about 90 meters below the K-Pg boundary and within the local range of Alamosaurus fossils, was dated to 69.0 plus or minus 0.9 million years old in 2010. Using this date, in correlation with a measured age from the underlying Aguja Formation and the likely location of the K-Pg boundary in the overlying Black Peaks Formation, the Alamosaurus fauna seems to have lasted from about 70-66 million years ago, with the earliest records of Alamosaurus near the base of the Javelina formation, and the latest just below the K-Pg boundary in the Black Peaks Formation. Skeletal elements of Alamosaurus are among the most common Late Cretaceous dinosaur fossils found in the United States Southwest and are now used to define the fauna of that time and place. In the south of Late Cretaceous North America, the transition from the Edmontonian to the Lancian is even more dramatic than it was in the north. Thomas M. Lehman describes it as "the abrupt reemergence of a fauna with a superficially 'Jurassic' aspect." These faunas are dominated by Alamosaurus and feature abundant Quetzalcoatlus in Texas. The Alamosaurus-Quetzalcoatlus association probably represent semi-arid inland plains. The appearance of Alamosaurus may have represented an immigration event from South America. Some taxa may have co-occurred on both continents, including Kritosaurus and Avisaurus. Alamosaurus appears and achieves dominance in its environment very abruptly, which might support the idea that it originated following an immigration event. Other scientists speculated that Alamosaurus was an immigrant from Asia. However, critics of the immigration hypothesis note that inhabitants of an upland environment like Alamosaurus are more likely to be endemic than coastal species, and tend to have less of an ability to cross bodies of water. Further, Early Cretaceous titanosaurs are already known, so North American potential ancestors for Alamosaurus already existed. Other contemporaneous dinosaurs from that part of the world include tyrannosaurs, smaller theropods, the hadrosaurs Edmontosaurus sp. and Kritosaurus sp., the ankylosaur Glyptodontopelta, and the ceratopsids Torosaurus utahensis and Ojoceratops fowleri.
Размеры тела в сравнении с человеком:
Ископаемые останки и реплики (1, 2, 3, 4):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| March 1st, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 07:52 pm [industrialterro] [Link] |
Ajkaceratops Айкацератопс (Ajkaceratops; от венгер. «Ajka» Айка — город в Венгрии и «ceratops» — рогатая морда) — род травоядных динозавров инфраотряда цератопсов, обитавший в меловом периоде (около 85 млн лет назад) на территории нынешней Европы. Типовой вид — Ajkaceratops kozmai. В длину взрослый Ajkaceratops достигал 1 метра. Окаменелые останки были найдены в 2009 году группой палеонтологов под руководством Аттилы Оси (Atilla Ösi) из Венгерской Академии Наук (Hungarian Academy of Sciences) на территории современной Венгрии в бокситовых рудниках Ихаркут близ города Айка и описаны в 2010 году. Описан по верхним и нижним частям клюва. Ученые предполагают, что A. kozmai начал своё путешествие из Азии и, двигаясь от острова к острову, постепенно уходил всё дальше на запад (в современную Европу). Ajkaceratops схож по строению с азиатскими видами цератопсов Bagaceratops и Magnirostris, что дает возможность предположительно отнести его к семейству протоцератопсидов, однако точную принадлежность установить трудно из-за недостаточного количества костей. Сравнение окаменелостей путем кладистического анализа говорит о том, что Ajkaceratops является цератопсидом, близким к Bagaceratops, но более примитивным, чем Zuniceratops и Ceratopsidae. На основе исследования окаменелостей Ajkaceratops Оси (2010) определил данного динозавра как базальный род клады Coronosauria. The holotype, cataloged as MTM V2009.192.1, consists only of a few skull fragments, including rostral bones, fused premaxillae, and maxillae fragments (the beak and jaw fragments). These fossils are kept in the Hungarian Natural History Museum, in Budapest. Although the fossils are fragmentary, the paper describing Ajkaceratops estimated a body length of 1 m (3.3 ft). Other material includes four predentary bones, cataloged as MTM V2009.193.1, V2009.194.1, V2009.195.1, and V2009.196.1; these are also believed to have belonged to Ajkaceratops, although they are proportionately smaller, and probably came from other individuals of the genus. The fossils of Ajkaceratops were discovered in the Csehbánya Formation, which is interpreted as "a floodplain and channel deposit formed by variegated clay, silt with interbedded grey and brown sand, and sandstone beds". This strata dates to the Santonian stage, around 86 to 84 mya (million years ago). Ajkaceratops shared its environment with other dinosaurs such as Rhabdodon, nodosaurid ankylosaurs, and theropods, as well as non-dinosaurian eusuchian crocodiles, azhdarchid pterosaurs, bothremydid turtles, teiid lizards, and enantiornithine birds. Bagaceratopidae is a family of neoceratopsian dinosaurs. It was named by Alifanov in 2003 but no definition has been proposed. Because of lacking of the definition, Bagaceratopidae was considered inactive by Paul Sereno in 2005. Alifanov in 2003 classified to this family four genera: Bagaceratops, Breviceratops, Lamaceratops and Platyceratops and in a publication from 2008 he included in Bagaceratopidae also Magnirostris and newly described genus Gobiceratops. Alifanov suggested also that Bagaceratopidae, unlike other neoceratopsian families, is of Paleoasiatic origin. Bagaceratopids existed during the late Cretaceous period, between about 85.8 and 70.6 million years ago. Фрагменты черепа небольшого динозавра, относившегося к инфраотряду цератопсов (Ceratopsia), были найдены в Венгрии. Три фрагмента палеонтологи под руководством Аттилы Оси (Attila Osi) из Венгерской академии наук (Hungarian Academy of Sciences in Budapest) нашли в бокситовом карьере возле города Айка (Ajka). Палеонтологи назвали нового динозавра в честь этого города - Ajkaceratops kozmai. Статья, посвященная открытию этого динозавра, опубликована в журнале Nature. О том, что цератопсы обитали в Европе, палеонтологи догадывались уже давно. Ведь первые представители этой группы возникли в Азии, а своего расцвета цератопсы достигли в Азии и Северной Америке. Попасть в Америку минуя Европу в конце юрского и в меловом периоде было практически невозможно. Поэтому, логично было предположить, что они расселялись как раз через Европу, переплывая с острова на остров или мигрируя в те эпохи, когда море отступало и острова сливались между собой. Но костей в Европе ранее не находили, поэтому вопрос оставался открытым до обнаружения айкацератопса. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, багацератопсиды, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, птицетазовые, цераподы, цератопсиды, цератопсы | ||||||||||||||||||||||||||||||||||||||||
| 06:57 pm [industrialterro] [Link] |
Agustinia Агустиния (Agustinia ligabuei) — вид ящеротазовых динозавров монотипного рода Agustinia из группы зауроподоморфов, живших в раннем меловом периоде (около 116—100 миллионов лет назад), на территории нынешней Южной Америки. Окаменелости были найдены в провинции Неукен (формация Lohan Cura) в Аргентине. Впервые описан палеонтологом Бонапарте в 1999 году. The name Agustinia honors the discoverer of the specimen, Agustin Martinelli. This dinosaur was originally named in a 1998 abstract written by famous Argentine paleontologist Jose Bonaparte. The original generic name was "Augustia", which, as it turned out, was already preoccupied by a beetle (see also: Megapnosaurus, Protognathosaurus). Bonaparte changed the name to Agustinia in a full paper published in 1999. There is one named species (A. ligabuei), which is named in honor of Dr. Giancarlo Ligabue, a philanthropist who provided financial support to the expedition which recovered the remains. Only fragmentary remains are known. These include fragments of vertebrae from the back, hips, and tail regions of the spinal column. Parts of the lower hind limb were also recovered: a fibula, tibia, and 5 metatarsals. A femur (thigh bone) was found at the site but was too fragmented to collect. Agustinia ligabuei was well known for its distinctive supposed armor plates, initially interpreted as a series of wide, vertical spikes and plates down the center of its back, somewhat like the unrelated Stegosaurus. However, further study of the poorly preserved fossil material showed that these "plates" were in fact more likely to be fragments of ribs and hip bones. Aside from the supposed armor, very little has been described of the anatomy of Agustinia. A fibula (lower leg bone) has been recovered that is about 3 feet (895 mm) long. When compared to the same bone in related dinosaurs, this indicates that Agustinia may have been about 50 feet (15 meters) long. However, the remains are incomplete and do not appear to have many distinctive characteristics which can be used to separate it from other sauropods. With the supposedly distinctive armor discredited, some authors have considered Agustinia a nomen dubium, based on remains not adequate to compare with related dinosaurs. Because of its supposedly unusual armor, Agustinia was originally assigned to its own family, Agustiniidae (Bonaparte, 1999). This family name has not come into wide acceptance. Agustinia is difficult to classify because of its fragmentary nature, and because it exhibits features of both diplodocoid and titanosaurian sauropods. What few distinctive features do exist in the only known specimen suggests it is a member of the clade Somphospondyli, but it is not possible to determine its relationships with any more accuracy beyond that. Somphospondyli — клада динозавров-зауроподов в составе более широкой группы Titanosauriformes. В 1998 году Пол Серено определил данную группу как состоящую из всех Titanosauriformes и более тесно связанную с Saltasaurus, чем с брахиозавром. В 2005 году Серено дал более точное определение: группа, состоящая из Saltasaurus loricatus и всех видов, более тесно связанных с Saltasaurus, чем с Brachiosaurus brancai. Название означает «губчатый позвонок» ссылаясь на синапоморфию группы. Somphospondyli по определению являются сестринской группой Brachiosauridae. Базальным членом группы вероятно является Erketu, найденный в 2006 году. Таксон по некоторым определениям является синонимом клады Titanosauria, хотя последняя не отвечает требованиям именования биологических таксонов согласно Международному кодексу филогенетических номенклатур. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, сомфоспондилы, титанозавроформы, ящеротазовые | ||||||||||||||||||||||||||||||||||||||||
| February 22nd, 2015 | |||||||||||||||||||||||||||||||||||||||||
| 07:13 pm [industrialterro] [Link] |
Agujaceratops Агухацератопс (Agujaceratops) — род птицетазовых динозавров из семейства цератопсидов, группы маргиноцефалов, живших в позднем меловом периоде (около 83—70 миллионов лет назад), на территории нынешней Северной Америки. Окаменелости были найдены в округе Брюстер (Brewster) штата Техас, формации Агуха (Aguja Formation). Впервые описан палеонтологами Лукасом (Lucas), Салливаном (Sullivan) и Хантом (Hunt) в 2006 году. Представлен одним видом — A. mariscalensis. Известен только один скелет. Первоначально Леманом (Lehman) в 1989 году был назван Chasmosaurus mariscalensis. Экземпляр был пронумерован P.37.3.086 UTEP. Однако позже, в 2006 году, был рекомбинирован в A. mariscalensis Лукасом и его коллегами. In 1938, three dinosaur bone beds were excavated, and ceratopsian material was collected from Big Bend National Park (Texas) by William Strain. This material was studied by Lehman in 1989 and named Chasmosaurus mariscalensis. It is known only from the holotype UTEP P.37.7.086 a partial adult skull which includes a braincase, left supraorbital horncore, left maxilla and a right dentary. Additional material was associated with the holotype, but not considered to be part of it. All specimens of Agujaceratops were collected from the lower part of the Upper Shale member of the Aguja Formation, dating to about 77 million years ago, in the Big Bend National Park, Brewster County. Subsequent analysis resulted in the taxon being put in its own genus. Agujaceratops was named by Spencer G. Lucas, Robert M. Sullivan and Adrian Hunt in 2006, and the type species is Agujaceratops mariscalensis. Agujaceratops is similar to both Pentaceratops and Chasmosaurus. Its short frill suggests it probably was not an ancestor of Pentaceratops. Агухацератопс был типичным представителем семейства цератопсид, жившим примерно 83-70 миллионов лет назад (кампанский ярус), на территории Северной Америки (США). Агухацератопс был длинной примерно в восемь метров, высотой почти до трех метров и как предполагают ученые весил около двух тонн. Динозавр имел черепной воротник схожий с воротником пентацератопса (родственного ему вида), который был более вертикально расположен и в длину немного короче. На тыльной стороне ободка воротника наблюдается достаточно глубокая вырезка. Череп массивный и тяжелый, в длину составлял примерно от метра до метра шестидесяти сантиметров. Над глазами находились два рога среднего размера, практически одинаковые и еще один небольшой рог располагался на носу. Рога были прямые, чуть загнутые кверху. Скорее всего, агухацератопс использовал свои внушительные рога для защиты при нападении хищников или при решении спорных вопросов во внутрисемейных конфликтах. По предположениям ученых-палеонтологов по морфологическим особенностям рогов и окраске данные динозавры могли определить возраст родственной особи, ее положение и авторитет в стаде. ( Read More )
Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, птицетазовые, хасмозаврины, цераподы, цератопсиды, цератопсы | ||||||||||||||||||||||||||||||||||||||||






















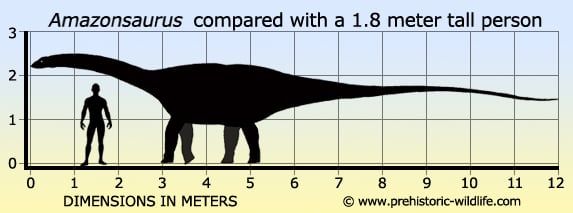












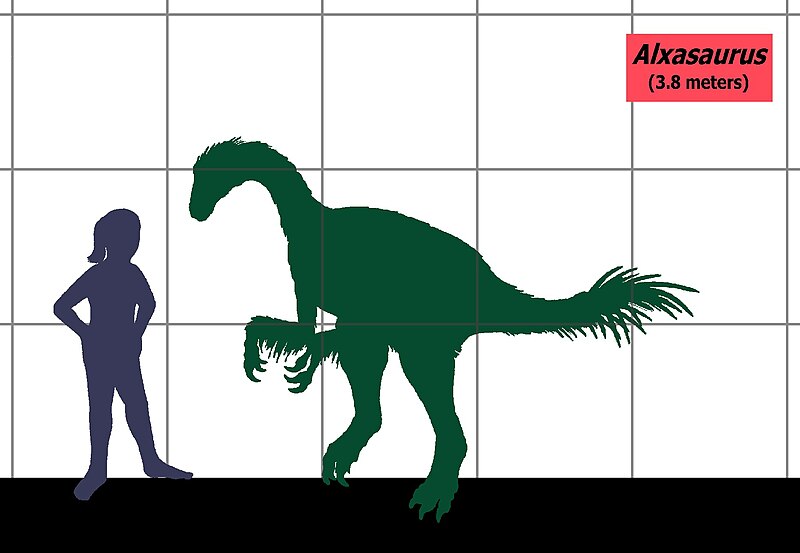





















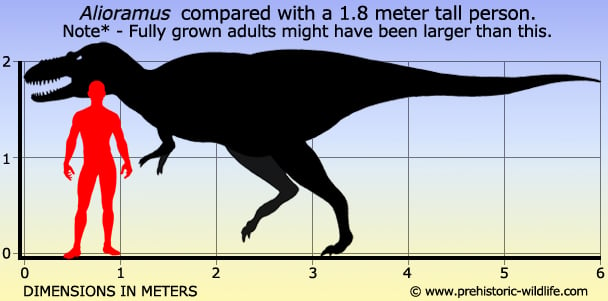








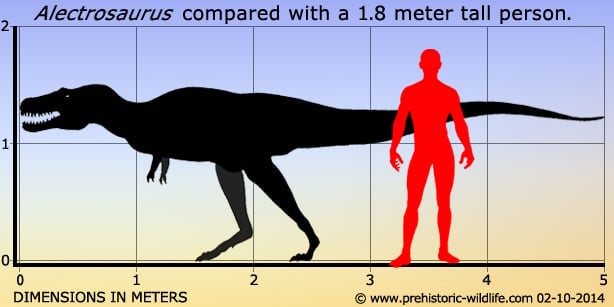





































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
_from_Mongolia.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}