[Recent Entries][Archive][Friends][User Info]
| May 29th, 2012 | |
|---|---|
| 08:51 pm [industrialterro] [Link] | Аллоза́вр (Allosaurus; греч. αλλος — «иной» или «странный», σαυρος — «ящер») — род хищных ящеротазовых динозавров подотряда тероподов, живших в юрском периоде (киммеридж — ранний титон) примерно 155—145 миллионов лет назад. Первые останки были изучены и классифицированы в 1877 году Гофниилом Чарлзом Маршем. Наряду с тираннозавром, аллозавр — самый популярный хищный динозавр. Он появлялся во множестве художественных фильмов, например, «Затерянный мир» 1925 года, или экранизации рассказа Р. Брэдбери «И грянул гром» 2005 года. Наиболее ярко и правдоподобно аллозавры были представлены в сериале BBC «Прогулки с динозаврами» и фильме «Баллада о Большом Але». Аллозавр был крупным двуногим хищником с большим черепом, оснащённым десятками больших острых зубов. Представители типового вида — А. фрагилис (A. fragilis) достигали в среднем 8,5 метра в длину, 3,5 метра в высоту и весили 2,5 - 3,5 тонны, хотя, на основании фрагментарных останков большего размера, можно предложить, что крупные особи могли достигать до 15 метров в длину, порядка 4 метров в высоту и массы около 4 - 5 тонн. Аллозавр передвигался на крупных и мощных задних лапах, в то время, как его передние конечности были относительно маленькими; на них имелось три крупных, загнутых когтя. Массивный череп уравновешивался длинным, тяжелым хвостом. Пока точное количество действительных видов неизвестно, но на сегодняшний день выделяются следующие: Allosaurus fragilis (fragilis — хрупкий) — типовой вид, описан О. Ч. Маршем в 1877 году. Поздняя юра (киммеридж — ранний титон) запада Северной Америки. Известен по огромному количеству образцов, включая полные скелеты различного размера, особей разного возраста из Колорадо, Юты, Вайоминга, Нью-Мексико. Описаны массовые захоронения в вязких асфальтовых или грязевых «ловушках для хищников», в Кливленд Лойд (40 особей). Длина до 8,5—12,3 метров, масса от 1 до 4 тонн, высота 3,5 метра. Иногда из этого вида выделяют ещё один — Allosaurus atrox (Creosaurus) — меньших размеров и с более низким черепом, из Вайоминга. Истинный статус креозавра неизвестен, но среди аллозавров вида Allosaurus fragilis наблюдаются две группы форм с преорбитальными рожками разной конфигурации. Возможно, это отражает половые различия. Недавно на основании находок почти полных скелетов в Юте и Вайоминге выделен вид Allosaurus jimmadseni, валидность которого признается не всеми авторами. Allosaurus europaeus — из позднего киммериджа — раннего титона Португалии. Чрезвычайно сходен с типовым видом, описан по неполному черепу в 2006 году. Allosaurus maximus — гигантский (весом более 2 тонн, длиной до 11—15 метров) аллозавр из киммериджа Оклахомы и Колорадо. Истинный статус неизвестен. Собственно Allosaurus maximus из Оклахомы часто выделяется в особый род Saurophaganax. К этому же виду иногда относят гигантского аллозавра эпантериаса (Epanterias amplexus) из Колорадо, которого обычно считают крупной особью типового вида. Кости аллозавров были найдены в позднеюрских отложениях Австралии, Африки и Северной Америки (Вайоминг, Юта, Колорадо). Неописанные виды аллозавров могут существовать в поздней юре США (например, «вайомингораптор» и другие), а также в поздней юре Китая и России. Знаменитый «Большой Ал», кстати, может принадлежать к неописанному ещё виду. Так называемый «карликовый полярный аллозавр» из раннего мела (альба) Австралии известен лишь по кости лодыжки и не может быть отнесен к роду Allosaurus. Африканский вид Allosaurus tendagurensis не может принадлежать к данному роду, но несомненно, принадлежит к аллозавридам. Вероятно, что в своё время крупный вид аллозавра был главным хищником и охотилися на крупных травоядных динозавров, таких как камарозавры и стегозавры, а, возможно, даже и на других хищников (например цератозавра). Есть доказательства (следы разных представителей одного вида в одном месте, массовые захоронения останков одного вида), того, что аллозавры охотились стаями, но некоторые палеонтологи считают, что они были слишком агрессивными для того, чтобы жить в стаях. Представители наиболее хорошо изученного вида A. fragilis в среднем достигали 8,5 метра в длину, наиболее крупные особи оцениваются в 9,7 метра и 2,3 веса тонны. В 1976 Джеймс Мэдсен изучил целый ряд скелетов разных размеров и видов и выяснил, что максимальная длина крупных видов достигала 12-13 метров. Точный вес аллозавра (как впрочем и всех динозавров)определить трудно. У аллозавра имелось шесть шейных позвонков, четырнадцать спинных и пять крестцовых. Число хвостовых позвонков неизвестно: Дж. Мэдсен считает, что их было не менее 50, а Григорий Пол, что не более 45. В позвонках аллозавра имелись сквозные отверстия. Подобные отверстия есть и у птиц: они помогают выталкивать воздух из воздушных мешков прямо через кожу, не тратя энергию на выдыхание из горла; что очень удобно при больших физических нагрузках (например, при полёте). Из этого следует, что аллозавр, вероятнее всего, интенсивно преследовал свою добычу — иначе объяснить наличие у него такого способа дыхания затруднительно. Возможно, что у аллозавра были дополнительные рёбра, как и у тираннозавра, но, возможно, это обломки костей или сильно фоссилизованная вилочковая кость, наличие которой у аллозавра было доказано в 1996 году. У некоторых особей аллозавров концы лобковых костей не соединенны; возможно, это помогало им лежать на земле; Джеймс Мэдсон полагает, что это помогало самкам откладывать яйца и являлось признаком полового диморфизма. Передние лапы аллозавра были короткими по сравнению с задними (у взрослых лишь около 35 % длины задних конечностей), на них имелись три пальца, которые кончались крупными, сильно изогнутыми когтями. Предплечья были несколько короче, чем плечи (соотношение длины плечевой и локтевой кости равнялось примерно 1:1.2); запястье равнялось по длине локтевой кости. Из трех пальцев на передней лапе средний был самым крупным и отличался от других количеством фаланг. Ноги аллозавра были приспособлены не столько для быстроты передвижения, сколько для устойчивости. На ноге аллозавра имелось три опорных пальца, и один, не использовавшийся при ходьбе. Также есть признаки того, что на задней лапе у аллозавра был и пятый рудиментарный палец. Череп аллозавра, по сравнению с черепами других тероподов, был небольшим. К примеру, череп тарбозавра был в два раза больше. Палеонтолог Г. С. Пол, изучив все известные черепа, пришёл к выводу, что самый большой достигал «всего» 845 мм. На каждой предчелюстной кости было пять зубов с D-образным сечением, а на каждой верхней челюстной кости, было от четырнадцати до семнадцати зубов, в зависимости от вида. На каждой нижней челюсти, было от четырнадцати до семнадцати зубов, чаще всего встречались черепа с шестнадцатью зубами на нижней челюсти. Зубы становились короче, уже и более изогнутыми по направлению к задней части черепа. Все зубы имели пилообразные края и легко сменялись после выпадения. Хорошо развитый шарнирный сустав между челюстями был сильно смещён к задней части черепа, что давало аллозавру возможность очень широко раскрывать пасть. Кроме того, в середине нижней челюсти было ещё одно сочленение, увеличивавшее эту возможность. На черепе были парные гребни, которые постепенно переходили в рожки. Эти рожки являлись увеличенными надбровными гребнями, которые у всех аллозавров были разными. Поверх костяного основания этих наростов, вероятно, шёл слой кератинового покрытия. Возможно эти гребни предназначались для защиты глаз от яркого солнечного света. Ранее считалось, что аллозавры бодались ими, но сейчас эта гипотеза отвергнута, так как эти рожки слишком хрупкие для такой цели. Внутри рожек, также могла располагаться сольная железа. Воздушные проходы у аллозавра были более развиты, чем у более примитивных тероподов, таких как цератозавр и маршозавр, благодаря чему у аллозавра было очень хорошо развито обоняние, а, возможно, имелся и вомероназальный орган. Лобные кости черепа была тонкими, возможно, для улучшения терморегуляции мозга. Аллозавр относился к семейству Аллозавридов из инфраотряда Карнозавров. Семейство Аллозавриды было предложено в 1878 году Гофониилом Чарльзом Маршем, но этот термин не употреблялся до 1970-х, а всех карнозавров относили в одно семейство Мегалозаврид. После публикации трудов Мадсена об аллозаврах, термин Аллозавриды стал использоваться многими палеонтологами. Как показывают исследования, представители семейства Аллозавридов обычно были крупнее Мегалозавридов. Очень близки к Аллозавридам, такие динозавры, как Индозавр, Пятницкозавр, Пиветеаузавр, Янхуанозавр, Акрокантозавр, Хилантаизавр, Компсозух, Стокеозавр и Сечуанозавр. Аллозавриды были одним из семейств, надсемейства Аллозавроидов к которым также относятся Кархародонтозавиды и Синорапториды. Ранее именно Аллозавроиды считались предками Тираннозавридов, но сейчас установлено, что это не так. К аллозавридам относится всего семь родов, но иногда называется больше, за счёт выделения Креозавра, Эпантериаса и Заврофагнакса в отдельные рода. В Кливленд Ллойде (расположенном вблизи Кливленда, штат Юта, США) был обнаружен так называемый «Карьер динозавров», в котором было найдено 44 скелета аллозавров (Allosaurus fragilis). В юрском периоде на этом месте было болото, в котором увяз гигантский брахиозавр и стая аллозавров набросилась на него, в результате чего многие из них оказались затянуты в трясину. В октябре 1965 года Кливленд Ллойд был объявлен национальным природным заповедником. Allosaurus was the most common large theropod in the vast tract of Western American fossil-bearing rock known as the Morrison Formation, accounting for 70 to 75% of theropod specimens, and as such was at the top trophic level of the Morrison food web. The Morrison Formation is interpreted as a semiarid environment with distinct wet and dry seasons, and flat floodplains. Vegetation varied from river-lining forests of conifers, tree ferns, and ferns (gallery forests), to fern savannas with occasional trees such as the Araucaria-like conifer Brachyphyllum. Allosaurus coexisted with fellow large theropods Ceratosaurus and Torvosaurus in both the United States and Portugal. The three appear to have had different ecological niches, based on anatomy and the location of fossils. Ceratosaurs and torvosaurs may have preferred to be active around waterways, and had lower, thinner bodies that would have given them an advantage in forest and underbrush terrains, whereas allosaurs were more compact, with longer legs, faster but less maneuverable, and seem to have preferred dry floodplains. Ceratosaurus, better known than Torvosaurus, differed noticeably from Allosaurus in functional anatomy by having a taller, narrower skull with large, broad teeth. Allosaurus was itself a potential food item to other carnivores, as illustrated by an Allosaurus pubic foot marked by the teeth of another theropod, probably Ceratosaurus or Torvosaurus. The location of the bone in the body (along the bottom margin of the torso and partially shielded by the legs), and the fact that it was among the most massive in the skeleton, indicates that the Allosaurus was being scavenged. Paleontologists accept Allosaurus as an active predator of large animals. There is dramatic evidence for allosaur attacks on Stegosaurus, including an Allosaurus tail vertebra with a partially healed puncture wound that fits a Stegosaurus tail spike, and a Stegosaurus neck plate with a U-shaped wound that correlates well with an Allosaurus snout. Sauropods seem to be likely candidates as both live prey and as objects of scavenging, based on the presence of scrapings on sauropod bones fitting allosaur teeth well and the presence of shed allosaur teeth with sauropod bones. However, as Gregory Paul noted in 1988, Allosaurus was probably not a predator of fully grown sauropods, unless it hunted in packs, as it had a modestly sized skull and relatively small teeth, and was greatly outweighed by contemporaneous sauropods. Another possibility is that it preferred to hunt juveniles instead of fully grown adults. Research in the 1990s and first decade of the 21st century may have found other solutions to this question. Robert T. Bakker, comparing Allosaurus to Cenozoic sabre-toothed carnivorous mammals, found similar adaptations, such as a reduction of jaw muscles and increase in neck muscles, and the ability to open the jaws extremely wide. Although Allosaurus did not have sabre teeth, Bakker suggested another mode of attack that would have used such neck and jaw adaptations: the short teeth in effect became small serrations on a saw-like cutting edge running the length of the upper jaw, which would have been driven into prey. This type of jaw would permit slashing attacks against much larger prey, with the goal of weakening the victim. Similar conclusions were drawn by another study using finite element analysis on an Allosaurus skull. According to their biomechanical analysis, the skull was very strong but had a relatively small bite force. By using jaw muscles only, it could produce a bite force of 805 to 2,148 N, less than the values for alligators (13,000 N), lions (4,167 N), and leopards (2,268 N), but the skull could withstand nearly 55,500 N of vertical force against the tooth row. The authors suggested that Allosaurus used its skull like a hatchet against prey, attacking open-mouthed, slashing flesh with its teeth, and tearing it away without splintering bones, unlike Tyrannosaurus, which is thought to have been capable of damaging bones. They also suggested that the architecture of the skull could have permitted the use of different strategies against different prey; the skull was light enough to allow attacks on smaller and more agile ornithopods, but strong enough for high-impact ambush attacks against larger prey like stegosaurids and sauropods. Their interpretations were challenged by other researchers, who found no modern analogues to a hatchet attack and considered it more likely that the skull was strong to compensate for its open construction when absorbing the stresses from struggling prey. The original authors noted that Allosaurus itself has no modern equivalent, that the tooth row is well-suited to such an attack, and that articulations in the skull cited by their detractors as problematic actually helped protect the palate and lessen stress. Another possibility for handling large prey is that theropods like Allosaurus were "flesh grazers" which could take bites of flesh out of living sauropods that were sufficient to sustain the predator so it would not have needed to expend the effort to kill the prey outright. This strategy would also potentially have allowed the prey to recover and be fed upon in a similar way later. An additional suggestion notes that ornithopods were the most common available dinosaurian prey, and that allosaurs may have subdued them by using an attack similar to that of modern big cats: grasping the prey with their forelimbs, and then making multiple bites on the throat to crush the trachea. This is compatible with other evidence that the forelimbs were strong and capable of restraining prey.
Карноза́вры (Carnosauria) — инфраотряд хищных динозавров, обитавших на Земле в юрском и меловом периодах около 168—65,5 миллионов лет назад. Изначально в эту группу определяли всех крупных хищных динозавров с короткой шеей (таких, как тираннозавр); в конце XX века, однако, выяснилось, что карнозавры имеют между собой мало общих признаков, кроме большого размера. Группа была переопределена и в настоящее время включает аллозавров и их сородичей, а тираннозавр и другие гиганты, не относящиеся к карнозаврам, были отнесены к другим таксонам (таким, как целурозавры или абелизавры). Среди карнозавров встречались и огромные динозавры, такие, как гиганотозавр, и сравнительно небольшие хищники (например, газозавр). В данном инфраотряде представлено несколько промежуточных таксонов: Аллозавроиды — надсемейство крупных динозавров, живших около 168—70 миллионов лет назад, в которое входит большая часть представителей инфраотряда, за исключение наиболее примитивных форм. Одним из самых ранних представителей является Poekilopleuron. Carcharodontosauria — группа хищных динозавров входящих в надсемейство аллозавроиды. Состоит из средних и крупных рептилий, живших в меловом периоде на территории Африки, Южной Америки, Австралии и Азии. Считается что представители данной группы появились около 130 миллионов лет назад, но самый старый известных представитель данной группы происходит из позднего Barremian. Последним представителем группы считается Orkoraptor, живший в маастрихте. Аллозаври́ды (Allosauridae) — семейство хищных ящеротазовых динозавров из группы теропод. Жили на территории нынешних Азии, Африки, Европы и Северной Америки около 155—145 миллионов лет назад. Семейство включает в себя Saurophaganax, Allosaurus, и мало изученного Epanterias, который может быть экземпляром Allosaurus. Впервые семейство было выделено американским палеонтологом Отниелом Чарльзом Маршем в 1878 году.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13):
Аллозавр (слева) отгоняет от добычи торвозавра (справа).
Аллозавр (справа) отгоняет от добычи стаю торвозавров (слева).
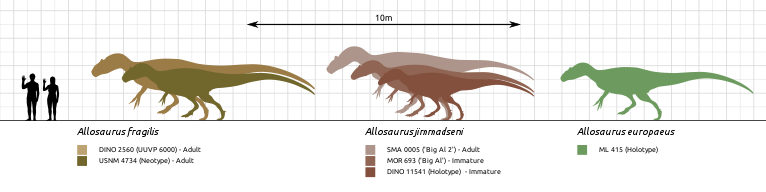
Размеры тела в сравнении с человеком:
Размеры тела в сравнении с другими гигантскими тероподами (закрашен фиолетовым):
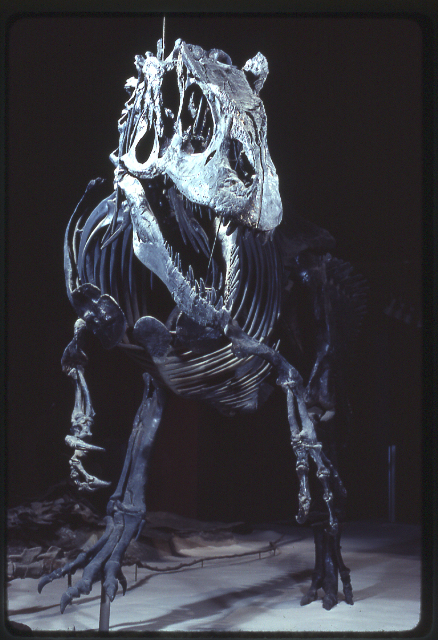
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9):
Tags: Вымершие рептилии, Юра, авеметатарзалии, аллозавриды, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, карнозавры, тероподы, ящеротазовые |






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}