[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 60 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| May 23rd, 2012 | |
|---|---|
| 08:22 pm [industrialterro] [Link] |
Puntanipterus Puntanipterus was a genus of dsungaripterid pterodactyloid pterosaur from the Upper Jurassic-Lower Cretaceous La Cruz Formation of San Luis, Argentina. The genus was in 1975 named by José Bonaparte and Teresa Sánchez. The type species is Puntanipterus globosus. The genus name refers to the Puntanos, the colloquial name for the inhabitants of the province of San Luis after the old name of their capital "San Luis de la Punta de los Venados", and combines this with a Latinized Greek pteron, "wing". The specific name means "spherical" in Latin, a reference to the form of the lower tibia. It is based on holotype PVL 3869 (earlier FML 3869) found in 1972, a 105 millimetres long tibiotarsus and seven centimetres long fibula; referred to it were a back vertebra and a wing and foot phalanx. The leg bones were described as similar to those of Pterodaustro (from slightly younger rocks), except for having an expanded spherical joint at the ankle and spiny processes on the side faces of the tibia at that end. Bonaparte in 1978 classified Puntanipterus as a member of the Pterodaustridae. The same year Peter Wellnhofer was more careful and limited his assessment to a Pterodactyloidea incertae sedis. In 1980 Peter Galton concluded it belonged to the Dsungaripteridae. It was still by many considered to be a dsungaripterid by the time Peter Wellnhofer published The Illustrated Encyclopedia of Pterosaurs (several editions in the 1990s). However, in the nineties several tibiae conforming to that of Puntanipterus were found in the same strata as Pterodaustro; a direct comparison is only impossible because more complete specimens of the latter are always very compressed, deforming the ankle morphology; but smaller fragments containing not-compressed ankles all have the build of a Puntanipterus tibiotarsus. This is by South American workers seen as a strong indication that both forms are identical. Glut reports a personal communication from Laura Codorniú and Luis Chiappe (2004) that Puntanipterus should be regarded as a junior synonym of Pterodaustro, but it remains to be seen if this will be supported in the future; it was not done in David Unwin's The Pterosaurs: From Deep Time, published in 2006 (he recognized it as a possibly valid species of uncertain relationships).
Tags: Вымершие рептилии, Юра, авеметатарзалии, аждархойды, архозавроморфы, архозавры, джунгариптероиды, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| 07:59 pm [industrialterro] [Link] |
Pterodactylus Pterodactylus ( The name derives from the Greek words pteron (πτερόν, meaning 'wing') and daktylos (δάκτυλος, meaning 'finger') and refers to the way in which the wing is supported by one large finger. Pterodactylus is known from over 27 fossil specimens, and though most of those are juveniles, many preserve complete skeletons. The discovery of several specimens with well-preserved soft tissue traces has allowed scientists to faithfully reconstruct the life appearance of Pterodactylus. Pterodactylus was a relatively small pterosaur, with an estimated adult wingspan of about 1.5 meters (5 ft) in P. antiquus. Other "species" were once thought to be smaller. However, these smaller specimens have been shown to represent juveniles of Pterodactylus, as well as its contemporary relatives Ctenochasma, Germanodactylus and Gnathosaurus. The skulls of adult Pterodactylus were long and narrow with about 90 large, conical teeth. The teeth extended back from the tips of both jaws, and became smaller farther away from the jaw tips (unlike some relatives, where teeth were absent in the upper jaw tip and were relatively uniform in size). The teeth extended farther back into the jaw than in close relatives, as some were present below the front of the nasoantorbital fenestra, the largest opening in the skull. Unlike related species, the skull and jaws were straight, not curved upwards. A small, hooked beak was present in the very tips of the jaws, with both upper and lower hook no larger than the teeth that surrounded them. The neck was long, and covered in long, bristle-like pycnofibres. A throat pouch extended from about the middle of the lower jaw to the upper part of the neck.ъ Pterodactylus, like related pterosaurs, had a crest on its skull composed mainly of soft tissues. In adult Pterodactylus, this crest extended between the back edge of the antorbital fenestra (the largest opening in the skull) and the back of the skull. The back of the crest extended upward into a backward-curving cone-shaped structure. The crest was composed mainly of long, hardened fibres (twisted together in a spiral pattern inside the conical part of the crest), and covered in scales. In at least one specimen of P. longicollum, the crest had a short bony base, also seen in related pterosaurs like Germanodactylus. Crests have only been found on large, fully adult specimens of Pterodactylus, indicating that this was a display structure and only developed when individuals reached maturity. The wings were long, and the wing membranes appear to have lacked the furry covering of pycnofibres present in some other pterosaurs (such as Pterorhynchus and Jeholopterus). The wing membrane extended between the fingers and toes as webbing, and a uropatagium (secondary membrane between the feet and tail) was present, as well as a propatagium (membrane between the wrist and shoulder). Both the finger and toe claws were covered in keratin sheaths that extended and curved into sharp hooks well beyond their bony cores. Like other pterosaurs (notably Rhamphorhynchus), Pterodactylus specimens can vary considerably based on age or level of maturity. Both the proportions of the limb bones, size and shape of the skull, and size and number of teeth changed as the animals grew. Historically, this has led to various growth stages (including growth stages of related pterosaurs) being mistaken for new species of Pterodactylus. Several detailed studies using various methods to measure growth curves among known specimens have demonstrated that there is actually only one valid Pterodactylus species, P. antiquus. The youngest immature Pterodactylus specimens have a small number of teeth (as few as 15), and the teeth have a relatively broad base. The teeth of older specimens are both narrower and more numerous (up to 90 teeth are present in some specimens). Pterodactylus specimens can be divided into two distinct year classes. In the first year class, the skulls are only 15-45mm in length. The second year class is characterized by skulls 55-95mm long, but still immature. These first two size groups were once classified as juveniles and adults of the species P. kochi, until further study showed that even the supposed "adults" were immature. A third year class is represented by specimens of the "traditional" P. antiquus, as well as a few isolated, large specimens once assigned to P. kochi that overlap P. antiquus in size. However, all specimens in this third year class also show sign of immaturity. Fully mature Pterodactylus specimens remain unknown, or may have been mistakenly classified as a different genus. The distinct year classes of Pterodactylus antiquus specimens show that this species, like the contemporary Rhamphorhynchus muensteri, likely bred seasonally and grew consistently during its lifetime. A new generation of 1st year class P. antiquus would have been produced seasonally, and reached 2nd-year size by the time the next generation hatched, creating distinct 'clumps' of similarly-sized and aged individuals in the fossil record. The smallest size class probably consisted of individuals that had just begun to fly and were less than one year old. The second year class represents individuals one to two years old, and the rare third year class is composed of specimens over two years old. This growth pattern is similar to modern crocodilians, rather than the rapid growth of modern birds. Comparisons between the scleral rings of Pterodactylus antiquus and modern birds and reptiles suggest that it may have been diurnal. This may also indicate niche partitioning with contemporary pterosaurs inferred to be nocturnal, such as Ctenochasma and Rhamphorhynchus. Numerous species have been assigned to Pterodactylus in the years since its discovery. In the first half of the nineteenth century any new pterosaur species would be named Pterodactylus, which thus became a typical "waste-basket taxon". Even after clearly different forms had later been given their own generic name, new species would be created from the very productive late Jurassic German sites, often based on only slightly different material. Around 1980, subsequent revisions by Peter Wellnhofer had reduced the number of recognized species to about half a dozen. Many species assigned to Pterodactylus had been based on juvenile specimens, and subsequently been recognized as immature individuals of other species or genera. By the 1990s it was understood that this was even true for part of the remaining species. P. elegans, for example, was found by numerous studies to be an immature Ctenochasma. Another species of Pterodactylus based on small, immature specimens is P. micronyx. However, it has been difficult to determine exactly of what genus and species P. micronyx might be the juvenile form. Stéphane Jouve, Christopher Bennett and others suggested that it probably belongs either to Gnathosaurus subulatus or one of the Ctenochasma species, but more data and study would be required to determine which one. The only well-known and well-supported species left were P. antiquus and P. kochi. However, most studies since the 1990s have found little reason to separate even these two, and have treated them as synonymous. In 1996, Bennett suggested that the differences between specimens of P. kochi and P. antiquus could be explained by differences in age. In a 2004 paper, Jouve used a different method of analysis and recovered the same result, showing that the "distinctive" features of P. kochi were age-related, and using mathematical comparison to show that the two forms are different growth stages of the same species. A special case is P. longicollum, named by von Meyer in 1854, based on a large specimen with a long neck and fewer teeth. Many researchers, including David Unwin, have found P. longicollum to be distinct from P. kochi and P. antiquus. Unwin found P. longicollum to be closer to Germanodactylus and therefore requiring a new genus name. It has sometimes been placed in the genus Diopecephalus because Harry Govier Seeley based this genus partly on the P. longicollum material. However, it was shown by Bennett that the type specimen later designated for Diopecephalus was a fossil belonging to P. kochi, and no longer thought to be separate from Pterodactylus. Diopecephalus is therefore a synonym of Pterodactylus, and as such is unavailable for use as a new genus for "P." longicollum.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Размеры тела в сравнении с человеком (серым показаны гипотетические размеры взрослой особи, зелёным - размеры найденных сеголетков):
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, ктенохазматоидеи, монофенестраты, птеродактили, птерозавры |
| May 22nd, 2012 | |
| 08:11 pm [industrialterro] [Link] |
Nesodactylus Nesodactylus was a genus of "rhamphorhynchoid" pterosaur from the Oxfordian-age Upper Jurassic Jagua Formation of Pinar del Río, western Cuba. Its remains were collected but not prepared by Barnum Brown in 1918, from rocks better known for their fossils of marine life. When seven black chalkstone blocks were prepared from 1966 by Richard Lund by dissolving the substrate in acid, this revealed the remains of a pterosaur. Ned Colbert described and named the genus in 1969. The type species is Nesodactylus hesperius. The genus name is derived from Greek nesos, "island" and daktylos, "finger", a reference to the island of Cuba and the typical wing finger of pterosaurs. The specific name means "western", from Greek hesperios. The genus is based on holotype AMNH 2000, a partial skeleton including a skull fragment, numerous vertebrae from all parts of the spine and tail, zygapophyses (interpreted by Colbert as ossified tendons) on the tail, the pectoral girdle and a very deeply keeled sternum, arms and partial hands, part of the pelvis, parts of both femorae, partial metatarsals, and ribs. The specimen was disarticulated but associated and not very compressed; during the preparation from the limestone with acid, the bones were not completely removed. Colbert found Nesodactylus to have had longer wings and more robust limbs and longer legs than related Rhamphorhynchus, although of a similar size and overall anatomy. He classified it as a rhamphorhynchid and more precisely as a member of the Rhamphorhynchinae. In 1977 James A. Jensen and John Ostrom by mistake referred to it as Nesodon (1977). Although there is little overlapping material with contemporaneous Cacibupteryx, the two are clearly different based on details of the elbow and quadrate. At least one recent review suggests it was a rhamphorhynchine, while another does not classify it.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 07:39 pm [industrialterro] [Link] |
Jeholopterus Jeholopterus was a small anurognathid pterosaur from the Daohugou Beds of northeastern China (of uncertain age, probably Middle or Late Jurassic), between 168 and 152 million years ago), preserved with hair and skin remains. The type species is based on holotype IVPP V12705, a nearly complete specimen from the Daohugou beds of Ningcheng County in the Neimongol (Inner Mongolia) Autonomous Region of China. The specimen is crushed into a slab and counterslab pair, so that parts of the specimen are preserved on one side of a split stone and some on the other. This includes exquisite preservation of carbonized skin fibers and, arguably, "hair" or "protofeathers." The fibers are preserved around the body of the specimen in a "halo." Wing tissue is preserved, though its extent is debatable, including the exact points of attachment to the legs (or if it attached to the legs at all). In 2009 Alexander Kellner published a study reporting the presence of three layers of fibres in the wing, allowing the animal to precisely adapt the wing profile. As an anurognathid, Jeholopterus shows the skull form typical for this group, being wider than it was long (28 mm), with a very broad mouth. Most teeth are small and peg-like, but some are longer and recurved. The neck was short with seven or eight cervical vertebrae. Twelve or thirteen dorsal vertebrae are present and three sacrals. There are five pairs of belly ribs. The tail vertebrae have not been preserved. The describers argue that Jeholopterus had a short tail, a feature seen in other anurognathids but unusual for "rhamphorhynchoid" (i.e. basal) pterosaurs that typically have a long tail. Wang et al. cited the presence of a fringe of hair in the region of the tail to infer the presence of a short tail. However, a subsequent study by Dalla Vecchia argued that gleaning any information about the tail is impossible, given that the tail is "totally absent" in the fossil. The wing bones are robust. The metacarpals are very short. A short pteroid, supporting a propatagium, is pointing towards the body. The hand claws are long and curved. The wings of Jeholopterus show evidence that they attached to the ankle, according to Wang et al.. They are relatively elongated with a wingspan of ninety centimetres. The legs are short but robust. The toes bear well-developed curved claws, but these are not as long as the hand claws. The fifth toe is elongated, according to the authors supporting a membrane between the legs, the uropatagium. Jeholopterus was by the authors assigned to the Anurognathidae. In 2003 a cladistic analysis by Kellner found it to be a member, together with Dendrorhynchoides and Batrachognathus of an anurognathid clade Asiaticognathidae. An analysis by Lü Junchang in 2006 resolved its position as being the sister taxon of Batrachognathus. Далее идёт пятиминутка острого, запущенного хумора: Though he never examined the fossil himself, advertising artist David Peters has popularized his idiosyncratic opinions about Jeholopterus and other pterosaurs widely on the internet. In general he finds and illustrates hosts of ornamental features and even multiple embryos although no other researchers have ever confirmed his findings. By manipulating downloaded images of Jeholopterus in the computer art program Photoshop, David Peters (2003) reported that he discovered an unusual suite of soft-tissue remains, including a horse-like tail Peters speculated may have been used as a fly sweeper/distractor, as well as a long fly lure (similar to that of the anglerfish) protruding from the head, and a fin or series of fins along the back. Peters also reported that he had found "rattlesnake-like fangs", and these, along with what he described as a rattle-snake-like mandible, buttressed palate, "surgically-sharp" unguals, robust limbs and other characters suggested that Jeholopterus was a vampire pterosaur adapted to plunging fangs into tough hide, then rotating the skull forward locking the fangs beneath the hide to improve adhesion. The small teeth of the lower jaw would not have penetrated but squeezed the wound like a pliers. Prominent pterosaur researcher Chris Bennett has described Peters' findings as "fantasy" and has vehemently denounced his methodology.
Tags: Вымершие рептилии, Юра, авеметатарзалии, анурогнатиды, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 07:26 pm [industrialterro] [Link] |
Gnathosaurus Gnathosaurus (meaning 'jaw lizard') is a genus of ctenochasmatid pterosaur known from a single species, G. subulatus, described in 1833. This pterosaur had an estimated wingspan of about 1.7 meters. The slender, 28-cm-long skull had up to 130 needle-like teeth arranged laterally around the spoon-shaped tip. Fragments of Gnathosaurus jaw were first discovered in 1832 in the Solnhofen limestones of Southern Germany but were mistaken for a piece of teleosaurid crocodile jaw, hence the synonym Crocodylus multidens. Only when a complete skull was found in 1951 this animal was found to have been a pterosaur. The teeth arranged in a spoon shape may have been used to strain water for small animals, although this is conjectural. Several paleontologists, such as Christopher Bennett, have suggested that a purported tiny Pterodactylus species, P. micronyx, is likely a juvenile of Gnathosaurus subulatus. Ctenochasmatoidea is a group of pterosaurs within the suborder Pterodactyloidea. The earliest known ctenochasmatoid remains have been found in the Stonesfield Slate formation (UK), which dates to the Bathonian stage of the Middle Jurassic, dated to about 166 million years ago.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, ктенохазматоидеи, монофенестраты, птеродактили, птерозавры |
| 07:02 pm [industrialterro] [Link] |
Germanodactylus Germanodactylus ("German finger") is a genus of dsungaripteroid pterodactyloid pterosaur from Late Jurassic-age rocks of Germany, including the Solnhofen limestone. Its specimens were long thought to pertain to Pterodactylus. Its head crest is a distinctive feature. Germanodactylus is described as being "raven-sized" in weight. G. cristatus had a 0.98 wingspan (3.2 ft) and a 13 centimeter long (5.1 in) skull, while G. rhamphastinus was somewhat larger, with a 1.08 meter wingspan (3.5 ft) and a skull 21 centimeters long (8.3 in). Germanodactylus is known for its head crest, which had a bony portion (a low ridge running up the midline of the skull) and a soft-tissue portion that more than doubled its height. The bony part does not go as far up the head in G. cristatus as in G. rhamphastinus. The soft-tissue portion was not known early on, being first described in 2002 by S. Christopher Bennett. It was probably composed of cornified epidermis. Germanodactylus is the first genus for which a soft-tissue component of the crest is known, but similar structures were probably widespread among pterosaurs. This genus is unspecialized compared to the pterosaurs of the Cretaceous, and has had varying placements in Pterosauria. Yang Zhongjian, who named the genus, gave it its own family Germanodactylidae. Bennett included the genus in the family Pterodactylidae, and Alexander W.A. Kellner found it to be related to Pterodactylus in his 2003 phylogenetic analysis. David M. Unwin, on the other hand, preferred to consider it a basal dsungaripteroid, a group that evolved into dedicated shellfish-eaters. G. cristatus is based on specimen BSP 1892.IV.1, from the Solnhofen limestone of Eichstätt, Germany. It was originally described by Plieninger in 1901 as a specimen of Pterodactylus kochi, and was given its current specific name by Carl Wiman in 1925, meaning "crested" in Latin. Yang Zhongjian determined that it deserved its own genus in 1964. Second species G. ramphastinus (in 1858 accidentally revised to rhamphastinus by Christian Erich Hermann von Meyer) was named as a distinct species long before G. cristatus, described by Johann Andreas Wagner in 1851 as a species of the now deprecated genus Ornithocephalus. The specific name refers to the toucan, ramphastinos in Greek. It is based on specimen BSP AS.I.745, a skeleton from the slightly younger Mörnsheimer Limestone of Daiting, Germany. Peter Wellnhofer added it to Germanodactylus in 1970, although Maisch and his coauthors have suggested that it deserves its own genus, "Daitingopterus" David M. Unwin has also referred miscellaneous limb bones and vertebrae from the somewhat older Kimmeridge Clay of Dorset, England to the genus; these finds at the time marked the earliest appearance of short-tailed pterosaurs in the fossil record. Bennett suggested in 1996 that Germanodactylus represented adults of Pterodactylus, but this has been rejected by further studies, including his own. Bennett's 2006 reappraisal of Germanodactylus found both species to be valid and included within the genus, with G. cristatus known from four specimens including two juveniles, and G. rhamphastinus from two specimens. The genus differs from other pterosaurs by a combination of characteristics including a sharply pointed jaw tip, 4-5 premaxillary teeth and 8-12 maxillary teeth per side of the upper jaw, robust maxillary teeth that, unlike in Pterodactylus, are not reduced in size farther from the tip of the jaw, a naso-antorbital fenestra twice the length of the eye socket, and various proportional differences. G. cristatus differs from G. rhampastinus by having no teeth in the tip of the jaw and fewer teeth (~13 in each side of the upper jaw and ~12 in the lower versus 16 upper and 15 lower on each side for G. rhamphastinus). Dsungaripteroidea is a group of pterosaurs within the suborder Pterodactyloidea. The earliest known fossils attributed to this group are from the Kimmeridgian-age Upper Jurassic Argiles d'Octeville Formation of France, dated to around 155 million years ago, and belonging to the species Normannognathus wellnhoferi. The last known dsungaripteroid species is Lonchognathosaurus acutirostris, from the Albian-age Lower Cretaceous Lianmuqin Formation of Xinjiang, China, about 112 million years ago.
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Юра, авеметатарзалии, аждархойды, архозавроморфы, архозавры, джунгариптероиды, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| May 17th, 2012 | |
| 07:46 pm [industrialterro] [Link] |
Dorygnathus Dorygnathus ("spear jaw") was a genus of pterosaur that lived in Europe during the Early Jurassic period, 180 million years ago when shallow seas flooded much of the continent. It had a short 1.5 meter (five feet) wingspan, and a relatively small triangular sternum, which is where its flight muscles attached. Its skull was long and its eye sockets were the largest opening therein. Large curved fangs that "intermeshed" when the jaws closed featured prominently at the front of the snout while smaller, straighter teeth lined the back. Having variable teeth, a condition called heterodonty, is rare in modern reptiles but more common in primitive pterosaurs. The heterodont dentition in Dorygnathus is consistent with a piscivorous (fish-eating) diet. The fifth digit on the hindlimbs of Dorygnathus was unusually long and oriented to the side. Its function is not certain, but the toe may have supported a membrane like those supported by its wing-fingers and pteroids. Dorygnathus was according to David Unwin related to the Late Jurassic pterosaur, Rhamphorhynchus and was a contemporary of Campylognathoides in Holzmaden and Ohmden. Dorygnathus in general has the build of a basal, i.e. non-pterodactyloid pterosaur: a short neck, a long tail and short metacarpals — although for a basal pterosaur the neck and metacarpals of Dorygnathus are again relatively long. The skull is elongated and pointed. The largest known cranium, that of specimen MBR 1920.16 prepared by Bernard Hauff in 1915 and eventually acquired by the Humboldt Museum in Berlin, has a length of sixteen centimetres. In the skull the eye socket forms the largest opening, larger than the fenestra antorbitalis that is clearly separated from the slit-like bony naris. No bony crest is visible on the rather straight top of the skull or snout. The lower jaws are thin at the back but deeper toward the front where they fuse into the symphysis ending in a toothless point after which the genus has been named. In MBR 1920.16, the mandibula as a whole has a length of 147 millimetres. In the lower jaws the first three pairs of teeth are very long, sharp and pointing outwards and forwards. They contrast with a row of eight or more upright-standing much smaller teeth that gradually diminish in size towards the back of the lower jaw. No such extreme contrast exists in the upper jaws, but the four teeth in the premaxilla are longer than the seven in the maxilla that again become smaller posteriorly. The total number of teeth is thus at least 44. The long upper and lower front teeth interlaced when the beak was closed; due to their extreme length they then projected considerably beyond the upper and lower margins of the head. According to Padian, eight cervical, fourteen dorsal, three or four sacral and twenty-seven or twenty-eight caudal vertebrae are present. The exceptional fourth sacral is the first of the normal caudal series. The number of caudals is not certain because their limits are obscured by long thread-like extensions, stiffening the tail. The cervical vertebrae are rather long and strongly built, their upper surface having a roughly square cross-section. They carry double-headed thin cervical ribs. The dorsal vertebrae are more rounded with flat spines; the first three or four carry ribs that contact the sternal ribs; the more posterior ribs contact the gastralia. The first five or six, rather short, caudal vertebrae form a flexible tail base. To the back the caudals grow longer and are immobilised by their intertwining extensions with a length of up to five vertebrae which together surround the caudals with a bony network, allowing the tail to function as a rudder. The breastbone is triangular and relatively small; Padian has suggested it may have been extended at its back with a cartilaginous tissue. It is connected to the coracoid which in older individuals is fused to the longer scapula forming a saddle-shaped shoulder joint. The humerus has a triangular deltopectoral crest and is pneumatised. The lower arm is 60% longer than the upper arm. From the five carpal bones in the wrist a short but robust pteroid points towards the neck, in the living animal a support for a flight membrane, the propatagium. The first three metacarpals are connected to three small fingers, equipped with short but strongly curved claws; the fourth to the wing finger, in which the second or third phalanx is the longest; the first or fourth the shortest. The wing finger supports the main flight membrane. In the pelvis, the ilium, ischium and pubis are fused. The ilium is elongated with a length of six vertebrae. The lower leg, in which the lower two thirds of the tibia and fibula of adult specimens are fused, is a third shorter than the thighbone, the head of which makes an angle of 45° with its shaft. The proximal tarsals are never fused in a separate astragalocalcaneum; a tibiotarsus is formed. The third metatarsal is the longest; the fifth is connected to a toe of which the second phalanx shows a 45° bend and has a blunt and broad end; it perhaps supported a membrane between the legs, a cruropatagium. In some specimens, soft parts have been preserved but these are rare and limited, providing little information. It is unknown whether the tail featured a vane on its end, as with Rhamphorhynchus. However, Ferdinand Broili reported the presence of hairs in specimen BSP 1938 I 49, an indication that Dorygnathus also had fur and an elevated metabolism, as is presently assumed for all pterosaurs. In 1971 Rupert Wild described and named a second species: Dorygnathus mistelgauensis, based on a specimen collected in a brick pit near the railway station of Mistelgau, to which the specific name refers, by teacher H. Herppich, who donated it to the private collection of Günther Eicken, a local amateur paleontologist at Bayreuth, where it still resides. As a result the exemplar has no official inventory number. The fossil comprises a shoulder-blade with wing, a partial leg, a rib and a caudal vertebra. Wild justified the creation of a new species name by referring to the great size, with an about 50% larger wingspan than with a typical specimen; the short lower leg and the long wing. Padian in 2008 pointed out that D. banthensis specimen MBR 1977.21, the largest then known, has with a wingspan of 169 centimetres an even larger size; that wing and lower leg proportions are rather variable in D. banthensis and that the geological age is comparable. He concluded that D. mistelgauensis is a subjective junior synonym of D. banthensis. Dorygnathus is commonly thought to have had a piscivorous way of living, catching fish or other slippery sea-creatures with its long teeth. This is confirmed by the fact that the fossils have been found in marine sediments, deposited in the seas of the European Archipelago. In these it is present together with the pterosaur Campylognathoides that however is much more rare. Very young juveniles of Dorygnathus are unknown, the smallest discovered specimen having a wingspan of sixty centimetres; perhaps they were unable to venture far over open sea. Padian concluded that Dorygnathus after a relatively fast growth in its early years, faster than any modern reptile of the same size, kept slowly growing after having reached sexual maturity, which would have resulted in exceptionally large individuals with a 1.7 metres wingspan. On land, Dorygnathus was probably not a good climber; its claws show no special adaptations for this type of locomotion. According to Padian, Dorygnathus, as a small pterosaur with a long tail, was well capable of bipedal movement, though its long metacarpals would make him better suited for a quadrupedal walk than most basal pterosaurs. Most researchers however, today assume quadrupedality for all pterosaurs.
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 06:23 pm [industrialterro] [Link] |
Dimorphodon Диморфодон (двуформозуб) — вымершее летающее пресмыкающееся, жившее от 200 до 180 миллионов лет назад. Размах крыльев до 2 м, длина от кончика головы до кончика хвоста — 120 см, размер головы — до 30 см. Диморфодон – один из древнейших летающих динозавров (птерозавров), обитавший в юрском периоде. Этот ящер имел мощные крылья, размах которых составлял около 2 метров. Длина его тела с учётом хвоста составляла 1,2 метра, а размер головы около 30 сантиметров. Вытянутая передняя часть головы напоминала клюв, но, в отличие от аналогичной части тела современных птиц, он был усажен огромным количеством острых, загнутых назад зубов. Как на задних конечностях, так и на концах крыльев располагались острые когти. Всё это говорит о том, что диморфодон был хищником, но, судя по небольшим размерам, питался он в основном рыбой и насекомыми. Вероятно, диморфодоны могли не только летать, но и лазать по деревьям и даже несколько неуклюже передвигаться по суше, используя при этом все 4 конечности. Вид этот изучен довольно слабо, так как на данный момент были найдены останки лишь одного экземпляра на территории Англии. Вполне возможно, что диморфодоны обитали не только в Европе. Dimorphodon had a large, bulky skull approximately 22 centimetres in length, whose weight was reduced by large openings separated from each other by thin bony partitions. Its structure, reminiscent of the supporting arches of a bridge, prompted Richard Owen to declare that, in far as achieving great strength from light-weight materials was concerned, no vertebra was more economically constructed; Owen saw the vertebrate skull as a combination of four vertebrae modified from the ideal type of the vertebra. The front of the upper jaw had four or five fang-like teeth followed by an indeterminate number of smaller teeth; the maxilla of all exemplars is damaged at the back. The lower jaw had five longer teeth and thirty to forty tiny, flattened pointed teeth, shaped like a lancet. Many depictions give it a speculative puffin-like 'beak' because of similarities between the two animals' skulls. The body structure of Dimorphodon displays many "primitive" characters, such as, according to Owen, a very small brain-pan and proportionally short wings. The first phalanx in its flight finger is only slightly longer than its lower arm. The neck was short but strong and flexible and may have had a membraneous pouch on the underside. The vertebrae had pneumatic foramina, openings through which the air sacks could reach the hollow interior. Dimorphodon had an adult body length of 1 metre (3.3 ft) long, with a 1.45 meter (4.6 ft) wingspan. The tail of Dimorphodon was long and consisted of thirty vertebrae. The first five or six were short and flexible but the remainder gradually increased in length and were stiffened by elongated vertebral processes. The terminal end of the tail may have borne a Rhamphorhynchus-like tail vane, although no soft tissues have yet been found of Dimorphodon to confirm this speculation Owen saw Dimorphodon as a quadruped. He speculated that the fifth toe supported a membrane between the tail and the legs and that the animal was therefore very ungainly on the ground. His rival Harry Govier Seeley however, propagating the view that pterosaurs were warm-blooded and active, argued that Dimorphodon was either an agile quadruped or even a running biped due to its relatively well developed hindlimbs and characteristics of its pelvis. This hypothesis was revived by Kevin Padian in the nineteen eighties. However, fossilised track remains of other pterosaurs (ichnites) show a quadrupedal gait while on the ground and these traces are all attributed to derived pterosaurs with a short fifth toe. Dimorphodon's was elongated, clawless, and oriented to the side. David Unwin has therefore argued that even Dimorphodon was a quadruped, a view confirmed by computer modelling by Sarah Sangster. Our knowledge of how Dimorphodon lived is limited. It perhaps mainly inhabited coastal regions and might have had a very varied diet. Buckland suggested it ate insects. Later it became common to depict it as a piscivore (fish eater), though Buckland's original idea is more well supported by biomechanical studies. Dimorphodon had an advanced jaw musculature specialized for a "snap and hold" method of feeding. The jaw could close extremely quickly but with relatively little force or tooth penetration. This, along with the short and high skull and longer, pointed front teeth suggest Dimorphodon was an insectivore, though it may have occasionally eaten small vertebrates and carrion as well. In 1870 Seeley assigned Dimorphodon its own family, the Dimorphodontidae, with Dimorphodon as the only member. It was suggested in 1991 by German paleontologist Peter Wellnhofer that Dimorphodon might be descended from the earlier European pterosaur Peteinosaurus. Later exact cladistic analyses are not in agreement. According to Unwin, Dimorphodon was related to, though probably not a descendant of, Peteinosaurus, both forming the clade Dimorphodontidae, the most basal group of the Macronychoptera and within it the sister group of the Caelidracones. This would mean that both dimorphodontid species would be the most basal pterosaurs known with the exception of Preondactylus. According to Alexander Kellner however, Dimorphodon is far less basal and not a close relative of Peteinosaurus.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15):
( Далее ) Ископаемые останки (1, 2, 3, 4, 5, 6):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, диморфодонтиды, птерозавры, рамфоринхоидеи |
| May 15th, 2012 | |
| 08:38 pm [industrialterro] [Link] |
Dermodactylus Dermodactylus (meaning "skin finger", from Greek derma and daktylos, in reference to pterosaur wings being skin membranes supported by the ring fingers) was a genus of pterodactyloid (general term for "short-tailed" pterosaur) pterosaur from the Kimmeridgian-Tithonian-age Upper Jurassic Morrison Formation of Wyoming, USA. It is based on a single partial bone, from the hand. Dermodactylus is based on YPM 2000, a distal right fourth metacarpal found by Samuel Wendell Williston at Como Bluff. This bone constituted at the time the oldest pterosaur remains found, recognized, and described from North America. Othniel Charles Marsh first named it as a species of Pterodactylus: P. montanus, the specific name meaning "from the mountains" in Latin, but soon changed his mind and gave it a new generic name. At the same time he assigned another wing bone, teeth, vertebrae, and a scapulacoracoid to it, but this material is probably too large to belong to the type individual. Its place within the Pterosauria is uncertain, beyond the Pterodactyloidea. The material it is based on is too meager for further classification (although Carpenter et al.. [2003] note that the shape of the bone's articular end means that it did not belong to an ornithocheirid, a type of short-tailed pterosaur that often had a head crest and/or large teeth), or for adding additional remains to the genus with any certainty, and so it is now regarded as a dubious pterodactyloid. It was not even mentioned in the most recent major popular work on pterosaurs. Marsh suggested it had a wingspan of 1.5-1.8 meters (5-6 feet), but this is including the material excluded by Peter Wellnhofer, who estimates the wingspan of the only known individual at 1 meter (3.28 feet). John Foster estimates its weight at 3.3 kilograms (7.3 pounds). It would probably have been a small aerial carnivore. Птерода́ктили (Pterodactyloidea, от греч. πτερόν — «крыло» и δάκτυλος — «палец») — подотряд вымерших рептилий отряда летающих ящеров (птерозавров), живших в юрском и меловом периодах. Высокоспециализированная группа, приспособившаяся к жизни в воздухе. Для птеродактилей характерен сильно удлинённый лёгкий череп. Зубы немногочисленные и мелкие; чаще они отсутствовали. Шейные позвонки вытянутые, без шейных рёбер. Крылья мощные, широкие; летательные пальцы складывающиеся. Хвост очень короткий. Кости голени сращенные. Размеры птеродактилей сильно варьировали — от мелких, величиной с воробья, до гигантских птеранодонов с размахом крыльев до 8 метров, орнитохейрусов и аждархид (кецалькоатль, арамбургиана) с размахом крыльев до 12 метров. Мелкие питались насекомыми, крупные — рыбой и др. водными животными. Останки птеродактилей известны из верхнеюрских и меловых отложений Западной Европы, Восточной Африки и обеих Америк, Австралии, в России — Поволжья. На берегах Волги впервые останки птеродактиля обнаружены в 2005 году. Самый крупный птеродактиль был обнаружен в Румынии в местечке Себеш уезда Алба; размах его крыльев - 16 м. Отряд включает в себя ряд семейств: Istiodactylidae — семейство, представители которого обитали в юрском и меловом периодах. Все находки данного семейства сделаны в северном полушарии - Северной Америке, Европе и Азии. В 2011 году был описан новый вид Gwawinapterus beardi отнесенный в данное семейство. Он был найден на территории Канады в меловых отложениях датируемых концом мелового периода (75 млн лет назад). Pteranodontidae семейство больших птерозавров мелового периода живших в Северной Америке и Европе. Данное семейство включает следующие роды: Bogolubovia, Nyctosaurus, Pteranodon, Ornithostoma, Muzquizopteryx. Останки Ornithostoma, являющегося самым древним представителем семейства, были найдены в Великобритании. Tapejaridae известны по находкам из Китая и Бразилии времен раннего мелового периода. Azhdarchidae (от Ajdarxo, имя дракона в персидской мифологии, полученных от старого персидского Azi Dahaka) представляет собой семейство птерозавров, известное, прежде всего, с конца мелового периода, хотя ряд изолированных позвонков принадлежавших животным, относящимся к этой группе, известны и из раннего мела (140 млн. лет назад). Включает в себя некоторых представителей, являющихся самыми большими из известных летающих животных известных науке. В 2003 году D. M. Unwin определил птеродактилей как группа включающая Pteranodon longiceps, Quetzalcoatlus northropi, их последний общий предок и все его потомки.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, птеродактили, птерозавры |
| 08:05 pm [industrialterro] [Link] |
Darwinopterus Darwinopterus modularis — ископаемый вид птерозавров юрского периода. Первоначально, при его описани, был единственным в составе рода Darwinopterus, названного в честь Чарльза Дарвина. Ископаемые отложения Юрского периода: Китай, Ляонин, Jianchang, Tiaojishan Formation.ъ Дарвиноптерус был относительно маленьким птерозавром с длинным хвостом. Череп относительно крупный, имеет длину от 14 до 19 см, крылья были длиной от 34 до 46 см. На черепе находится низкий длинный гребень, похожий на таковой же у Noripterus и у базального рода Germanodactylus или других представителей семейства Dsungaripteroidea. В челюстях пятнадцать пар острых хорошо отделённых друг от друга зубов, постепенно увеличивающихся в направлении к передней части головы. Их форма говорит о функции удержания добычи. Голотип (с номером экземпляра ZMNH M8782; Zhejiang Museum of Natural History, Hanzhou, Zhejiang Province, Китай) — это хорошо сохранившийся скелет с черепом, челюстями, почти полным позвоночником, частично сохраненной грудиной, плечевым поясом, частично сохранившимися левой передней конечностью (крылом) и частями задних ног. Скелет передан в Естественноисторический Музей города Ханчжоу, провинция Чжэцзян. Второй экземпляр (YH-2000, учтённый при описании вида) частично сохранился: у него отсутствуют лишь части черепа, грудина и несколько костей пальцев. Он сохраняется в Yizhou Museum (Yixian, Yi) в провинции Ляонин. Авторами описания стали три палеонтолога из Института геологии Китайской Академии геологических наук (Junchang Lü, Yongqing Liu, Qiang Ji), сотрудник музея Zhejiang Museum of Natural History (Xingsheng Jin) и английский палеонтолог Дэвид Унвин (David M. Unwin; School of Museum Studies, University of Leicester, Лестер). Перовоначально был единственным видом рода Darwinopterus Lü et al., 2010 (позже было описано ещё два вида). Разделяет признаки характерные как для длиннохвостых, так и для короткохвостых птерозавров, поэтому описан как промежуточная между ними ископаемая группа (в составе таксона Breviquartossa Unwin, 2003; Monofenestrata). Darwinopterus linglongtaensis был описан в 2010 году китайским ученым Ван Сяолинем в соавторстве с другими исследователями. Обозначение вида относится к местности Linglongta. Голотип IVPP V16049 состоит из почти полного скелета молодого животного. Вид отличается от D. modularis в основном более высоким черепом и короткими тупыми зубами. Череп имеет длину 119,2 мм, бедренная кость равна десяти сантиметрам. Darwinopterus robustodens был описан в 2011 году китайскими исследователями Lü Junchang, Xu Li, Chang Huali и Zhang Xingliao. Обозначение вида означает «с надежными зубами». Вид был изучен на основе хорошо сохранившегося образца 41HIII-0309A, который отличается наличием больших зубов. В соответствии с исследованиями, зубы были адаптированы к поеданию жуков, которые обладали плотными щитками. Описание нового вида подняло вопрос о том, какое положение занимает Darwinopterus в родословной птерозавров. Точный кладистический анализ показал, что строение головы и шеи близко к представителям группы Pterodactyloidea. Но также данный род разделяет признаки группы Rhamphorhynchidae в составе клады Breviquartossa. Таким образом он был определён, как сестринский таксон по отношению к группе Pterodactyloidea. Для их объединения была выделена новая клада Monofenestrata Lü et al., 2010. Wukongopteridae — семейство вымерших птерозавров. Включает 5 видов из 4 родов. Обнаружены в Китае в формациях Tiaojishan и/или Daohugou Beds, которые относят с средней Юре. Впервые семейство Wukongopteridae было выделено в 2009 году китайскими палеонтологами (Wang et al. 2009), но без чётких диагностических признаков. Объём группы Wukongopteridae был очерчен в 2010 году (Wang et al. 2010) в составе видов Wukongopterus lii, Kunpengopterus sinensis, Darwinopterus modularis, Darwinopterus linglongtaensis и Changchengopterus pani. Представители семейства разделяют как примитивные, так и продвинутые черты птерозавров. Строение головы и шеи близко к представителям группы Pterodactyloidea, а другими признаками (длинным хвостом) они похожи на группу Rhamphorhynchidae. Палеонтологи из Китая и Великобритании сообщили об открытии нового вида птерозавров. В наше время обнаружением очередного, неизвестного ранее вида вымерших рептилий никого не удивишь, но это открытие, судя по всему, будет иметь огромное значение не только для понимания эволюции летающих ящеров - птерозавров, но и для понимания всего эволюционного процесса. Существо, удивительным образом сочетающее в себе признаки более древней и более молодой групп птерозавров палеонтологи назвали Darwinopterus modularis. Darwinopterus – в честь Чарльза Дарвина, ведь в 2009 году исполнилось 200 лет со дня рождения великого ученого и 150 лет со дня публикации его фундаментального труда, который принято коротко называть "Происхождение видов". Да и обнаружение "промежуточного звена" между двумя группа птерозавров является очевидным подтверждением правоты эволюционной теории, основоположником которой является Дарвин. Видовое название – modularis, палеонтологи дали птерозавру за его уникальное строение – он как бы собран из различных модулей. Именно в этом и заключается самая интересная часть открытия. Darwinopterus modularis по строению черепа и шейного отдела позвоночника – типичный птеродактиль, а вот по всему остальному – типичный рамфоринх. Darwinopterus modularis был относительно небольшим птерозавром. Длина его черепа составляла около 20 сантиметров, общая длина от кончика морды до кончика длинного хвоста не превышала 1-1,2 метра. Загнутые назад мощные острые зубы указывают на то, что дарвиноптерус был хищником. Судя по всему, это был экологический аналог ястреба – летающий хищник, охотящийся на небольшую добычу преимущественно в воздухе. Причем, охотился он не на насекомых, у насекомоядных рептилий зубы другие. Его добычей могли быть другие птерозавры, осваивающие воздух оперенные динозавры, предки птиц, возможно, планирующие между деревьями рептилии и даже млекопитающие. ( Далее )
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, вуконгоптериды, диапсиды, монофенестраты, птерозавры |
| May 9th, 2012 | |
| 04:12 pm [industrialterro] [Link] |
Campylognathoides Campylognathoides ("curved jaw", Strand 1928) was a genus of "rhamphorhynchoid" pterosaur, discovered in the Württemberg Lias deposits, the first specimen consisting of wing fragments. Further better preserved specimens were found in the Holzmaden shale and it was based on these specimens that Felix Plieninger erected a new genus. Compared to its contemporary from the same layers Dorygnathus, the snout on this genus is relatively short, though the skull is still in general elongated, be it much lighter built. The large eye sockets, placed low in the skull above a narrow jugal, have caused some researchers to speculate that Campylognathoides had especially acute vision, or possibly even a nocturnal lifestyle. The back of the skull is relatively high and flat, with a sudden downturn just in front of the eyes. The snout ends in a slender point curving a bit upwards at its very end. A large part of the snout is occupied by long bony nares. Below them a small triangular skull opening, the fenestra antorbitalis is present. Reflecting the more shallow snout, the teeth of Campylognathoides are also short and not at all laniaries or fang-like as in the markedly heterodont Dorygnathus. They are conical and recurved but have a broad base with the point bevelled off from the inside forming a sharp and strong cutting surface. In the upper jaw there are four rather widely spaced teeth in the praemaxilla gradually increasing in size from the front to the back; the fourth pair of teeth is the largest. Behind them are ten smaller teeth in the maxilla, gradually decreasing posteriorely. In the lower jaw there are twelve to fourteen teeth present in C. liasicus, sixteen to nineteen in C. zitteli. The largest total number is thus 66. According to a study by Kevin Padian there are eight cervical vertebrae, fourteen dorsals, four or five sacrals and up to 38 caudal vertebrae. The tail base is flexible with about six short vertebrae; behind them the caudals elongate and are stiffened by very long extensions allowing the tail to function as a rudder. The sternum of Campylognathoides was a rather large rectangular plate of bone with a short forward-facing crest called a cristospina. The upper arm is short but robust with a square deltopectoral crest. The lower arm too is short but wing length is considerable due to the hand, which has short metacarpals but a very long wing finger for a basal pterosaur, of which the second phalanx is the largest. The pteroid is short and robust. The pelvis is not very well known. A fossil collector found a well preserved Campylognathoides hip in a Braunschweig shale quarry in 1986. This pelvis, BSP 1985 I 87, proved to be scientifically significant because the hip socket was according to Peter Wellnhofer in an upward lateral position, preventing the animal from being able to orient its legs erectly like in dinosaurs, birds and mammals. This would prove that Campylognathoides was not well able to walk on its hind legs but must have walked quadrupedally. This gait posture has been confirmed in other "rhamphorhynchoids" (i.e. basal pterosaurs) as well. However, Padian in 2009 concluded the opposite, stating that an erected position was necessary to place the feet on the ground and that, though a quadrupedal gait was possible, a bipedal way of locomotion was a precondition for a fast gait. This subject remains highly controversial. The leg is rather short and the feet are small. The fifth toe, often interpreted as carrying a membrane between the legs, is exceptionally short for a basal pterosaur. Plieninger in his later publications assigned Campylognathus to the "Rhamphorhynchoidea". As this suborder is a paraphyletic assemblage of not specially related basal pterosaurs, this classification merely states the negative fact that it was not a pterodactyloid. A positive determination was first attempted by Baron Franz Nopcsa who in 1928 assigned the genus to the subfamily Rhamphorhynchinae within the family Rhamphorhynchidae. After a period in which very little work was done on pterosaur systematics, in 1967 Oskar Kuhn placed Campylognathoides in its own subfamily within the Rhamphorhynchidae, the Campylognathoidinae. However, in 1974 Peter Wellnhofer concluded that it was placed in a more basal position in the phylogenetic tree, below the Rhamphorhynchidae. In the early twenty-first century this was confirmed by the first extensive exact cladistic analyses. In 2003 both David Unwin and Alexander Kellner introduced a clade Campylognathoididae; within Unwin's terminology this clade is the sister clade of the Breviquartossa within the Lonchognatha; applying Kellner's terminology it is the most basal off-shoot within the Novialoidea. There is no material difference between the two positions. According to the analyses Campylognathoides would be closely related to Eudimorphodon, to which it is similar in skull, sternum and humerus form. This was confirmed by Padian in 2009, though Padian also pointed out several basal features present in Eudimorphodon but lacking in Campylognathoides. In 2010 an analysis was published by Brian Andres showing that Eudimorphodon together with Austriadactylus formed a very basal clade, leaving Campylognathoides as the only known member of the Campylognathoididae. Traditionally a piscivore lifestyle is attributed to Campylognathoides, as to most pterosaurs; in this case supported by the provenance of the finds from marine sediments and the very long wings. Padian however, has suggested that, in view of the stout short teeth, ideal for delivering a piercing bite, the form might well have been a predator of small terrestrial animals instead. The niche of specialised fish eater would then have been filled by Dorygnathus which is five times as common in the layers. The area of the fossil sites was in the early Jurassic located to the northwest of a large island , the Massif of Bohemia, situated in a shallow gulf of the Tethys Sea.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 04:01 pm [industrialterro] [Link] |
Batrachognathus Batrachognathus is an extinct genus of "rhamphorhynchoid" pterosaur from the Late Jurassic (Oxfordian - Kimmeridgian) Karabastau Svita of the central Asian republic of Kazakhstan. The genus was named in 1948 by the Russian paleontologist Anatoly Nicolaevich Ryabinin. The type species is Batrachognathus volans. The genus name is derived from Greek batrakhos, "frog" and gnathos, "jaw", in reference to the short wide head. The specific epithet means "flying" in Latin. Three fossils have been found in a lacustrine sediment in the North-West Tien Shan foothills of the Karatau Mountains. In the Jurassic this area had some similarities in habitat to the Solnhofen lagoon deposits in Bavaria, Germany. The genus is based on holotype PIN 52-2, an incomplete and disarticulated skeleton consisting of skull fragments, jaws, vertebrae, ribs, legs and wing bones. The skull of 48 mm long is high, short and broad. The upper jaws have in total 22 or 24 recurved conical teeth; with the lower jaws they make a short and very wide mouth. The animal is not preserved with a tail. Whether it had one is debatable; usually it is assumed a short tail was present. The wingspan has been estimated at 50 cm; David Unwin in 2000 gave a higher estimate of 75 cm. Batrachognathus was assigned to the Anurognathidae, as a relative of Anurognathus. In 2003 it was joined with the Asian anurognathids Dendrorhynchoides and Jeholopterus in a clade Asiaticognathidae by Alexander Kellner. According to an analysis in 2006 by Lü Junchang Batrachognathus and Jeholopterus were sister taxa. Like all anurognathids Batrachognathus is assumed to have been an insectivore, catching insects on the wing with its broad mouth.
Tags: Вымершие рептилии, Юра, авеметатарзалии, анурогнатиды, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 03:23 pm [industrialterro] [Link] |
Anurognathus Анурогнат (Anurognathus от греч. αν — без, греч. оυρα — хвост и греч. γναθος — челюсти) — род малых птерозавров семейства Anurognathidae, представители которого жили около 150 миллионов лет назад в конце юрского периода. Род был описан и назван Людвигом Дёдерляйном (Ludwig Döderlein) в 1923 году. Типовым видом является Anurognathus ammoni. Представители рода отличаются необычной внешностью по отношению к другим птерозаврам подотряда Рамфоринхоиды. Видовое название ammoni дано в честь баварского геолога Людвига фон Аммона, который собрал коллекцию ископаемых, в которой был и Anurognathus ammoni. Анурогнат означает буквально «бесхвостый» или «беззубый» — что ж, вполне понятная, хотя и не совсем точная характеристика применительно к этим необычным птерозаврам. Судить об анурогнатах ученым остаётся только по останкам одной-единственной особи, обнаруженной в Германии в 20-е годы XX века. Хвост у него и правда походил на обрубок, голова была маленькая и тупорылая, а в пасти имелось всего несколько мелких зубов. Зато анурогнаты обладали хорошо развитыми конечностями. Впрочем, наличие таких признаков объясняется тем, что найденные останки принадлежали молодой особи, — у взрослого животного они наверняка были бы другими. Кроме того, анурогнаты, возможно, принадлежали к легким охотникам, и добычей им служили стрекозы и прочие насекомые, которых они отлавливали, прыгая со спин динозавров, используя их как живые трамплины. В размахе крыльев достигали 50 см. Anurognathus had a short head with pin-like teeth for catching insects and although it traditionally is ascribed to the long-tailed pterosaur group "Rhamphorhynchoidea", its tail was comparatively short, allowing it more maneuverability for hunting. According to Döderlein the reduced tail of Anurognathus was similar to the pygostyle of modern birds. Its more typical "rhamphorhynchoid" characters include its elongated fifth toe and short metacarpals and neck. With an estimated wingspan of fifty centimetres (20 inches) and a nine centimetre long body (skull included), its weight was limited: in 2008 Mark Paul Witton estimated a mass of forty grammes for a specimen with a 35 centimetre wingspan. The holotype was redescribed by Peter Wellnhofer in 1975. Later a second, smaller, specimen was found, probably of a subadult individual. Its slab and counterslab are separated and both were sold to private collections; neither has an official registration. It was described by S. Christopher Bennet in 2007. This second exemplar is much more complete and better articulated. It shows impressions of a large part of the flight membrane and under UV-light remains of the muscles of the thigh and arm become visible. It provided new information on many points of the anatomy. The skull was shown to have been very short and broad, wider than long. It transpired that Wellnhofer had incorrectly reconstructed the skull in 1975, mistaking the large eye sockets for the fenestrae antorbitales, skull openings that in most pterosaurs are larger than the orbits but in Anurognathus are small and together with the nostrils placed at the front of the flat snout. The eyes pointed forwards to a degree, providing some binocular vision. Most of the skull consisted of bone struts. The presumed pygostyle was absent; investigating the real nine tail vertebrae instead of impressions showed that they were unfused, though very reduced. The wing finger lacked the fourth phalanx. According to Bennett a membrane, visible near the shin, showed that the wing contacted the ankle and was thus rather short and broad. Bennett also restudied the holotype, interpreting bumps on the jaws as an indication that hairs forming a protruding bristle were present on the snout. The Anurognathidae were a group of small pterosaurs, with short tails or tailless, that lived in Europe and Asia during the Jurassic and early Cretaceous periods. Four genera are known: Anurognathus, from the Late Jurassic of Germany, Jeholopterus, from the Middle or Late Jurassic of China, Dendrorhynchoides, from the Early Cretaceous of China, and Batrachognathus, from the Late Jurassic of Kazakhstan. Bennett (2007) claimed that the holotype of Mesadactylus, BYU 2024, a synsacrum, belonged to an Anurognathid. Mesadactylus is from the Late Jurassic Morrison Formation of the USA. Indeterminate Anurognathid remains have also been reported from the Middle Jurassic Bakhar Svita of Mongolia. A family Anurognathidae was named in 1928 by Franz Nopcsa von Felső-Szilvás (as the subfamily Anurognathinae) with Anurognathus as the type genus. The family name Anurognathidae was first used by Oskar Kuhn in 1967. Both Alexander Kellner and David Unwin in 2003 defined the group as a node clade: the last common ancestor of Anurognathus and Batrachognathus and all its descendants. The phylogeny of the Anurognathidae is uncertain. Some analyses, as those of Kellner, place them very basal in the pterosaur tree. However, they do have some characteristics in common with the derived Pterodactyloidea, such as the short and fused tail bones. In 2010 an analysis by Brian Andres indicated the Anurognathidae and the Pterodactyloidea were sister taxa. This conforms better to the fossil record because no early anurognathids are known and would require a ghost lineage of over sixty million years.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10):
( Далее )
Tags: Вымершие рептилии, Юра, авеметатарзалии, анурогнатиды, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 03:07 pm [industrialterro] [Link] |
Angustinaripterus Angustinaripterus was a basal pterosaur, belonging to the Breviquartossa, and discovered at Dashanpu near Zigong in the Szechuan province of China. Angustinaripterus was named in 1983 by He Xinlu. The type species is Angustinaripterus longicephalus. The genus name is derived from Latin angustus, "narrow" and naris, "nostril", combined with Latinized Greek pteron, "wing". The specific name is derived from Latin longus, "long", and Greek kephale, "head". The holotype, ZDM T8001, is a single skull with lower jaws, found in 1981 by researchers from the Zigong Historical Museum of the Salt Industry, in the Xiaximiao Formation (Bathonian). The skull, of which the left side is severely damaged, is very elongated and flat. The back part is missing; in its conserved state it has a length of 192 millimetres; the total length in a complete state was estimated at 201 millimetres. On its top is a low crest, two to three millimetres high. The nares are long, slit-like and positioned above and in front of the large skull openings, the fenestrae antorbitales, with which they are not confluent. Of the jaws, which are very straight, the front part is lacking. There are six pairs of teeth in the maxillae and three pairs in the praemaxillae. In the mandible there are at least ten pairs of teeth, perhaps twelve. The back teeth are small, the front teeth are very long, robust and curved, pointing moderately forwards. At the front they form a large, intermeshing "prey grab", that may have been used to snatch fish from the water surface. The teeth of Angustinaripterus resemble those of Dorygnathus. He placed Angustinaripterus into the Rhamphorhynchidae. Because of the derived morphology and the large geographical distance with comparable European forms He also created a special subfamily Angustinaripterinae, of which Angustinaripterus itself is the only known member; because of this redundancy the concept is rarely used. He concluded that Angustinaripterus was directly related to the Scaphognathinae. David Unwin however, considers it a member of the other rhamphorhynchid subgroup: the Rhamphorhynchinae. Peter Wellnhofer in 1991, assuming the skull length was 16.5 centimetres, estimated the wingspan at 1.6 metres.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхиды, рамфоринхоидеи |
| March 10th, 2012 | |
| 12:42 pm [industrialterro] [Link] |
Preondactylus Preondactylus is a genus of long-tailed pterosaur from the Late Triassic that inhabited what is now Italy. It was discovered by Nando Buffarini in 1982 near Udine in the Preone valley of the Italian Alps. Preondactylus had single cusp teeth, meaning they had one point on each tooth. Its diet either consisted of fish, insects or both, but there is still debate going on as the tooth structure could indicate either diet (or both). It had short wings, whose total span was only 45 cm (18 in), and relatively long legs. The short wings were a "primitive" feature for pterosaurs, but Preondactylus was a fully developed flier. When Buffarini first discovered Preondactylus, the thin slab of bituminous, dolomitic limestone containing the fossil was accidentally broken into pieces while being extracted. After reassembly the rock was cleaned with water by him and his wife and the marl and in it the bone was washed away and lost. All that was left was a negative imprint on the stone, of which a silicon rubber cast was made to allow for subsequent study of the otherwise lost remains. Most of the skeleton is known, but the posterior portions of the skull have not been preserved. This first specimen is the holotype: MFSN-1891. A second, disarticulated specimen, MFSN-1770, was found at the same locale in 1984 about 150-200 meters deeper into the strata than the original find. The second specimen appears to have been preserved in the gastric pellet of a predatory fish, which had consumed the pterosaur and vomited up the indigestible pieces that would later fossilize. More detailed knowledge of the variability of Triassic pterosaurs has made the identification of this specimen as Preondactylus uncertain. A third specimen is MFSN 25161, a partial skull, lacking the lower jaws. The species was described and named by Rupert Wild in 1984. The genus name refers to Preone, the specific name honours Buffarini. Rupert classified the new species within Rhamphorhynchidae, of which group very old species are known such as Dorygnathus, but soon it was understood the form was much more basal. A cladistic analysis by David Unwin found Preondactylus as the most basal pterosaur, and the species was accordingly used by him for a node clade definition of the clade Pterosauria. Other analyses however, have found a somewhat more derived position for Preondactylus.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 12:23 pm [industrialterro] [Link] |
Peteinosaurus Peteinosaurus (pronounced /pɛˌtaɪnəˈsɔrəs/ or /pɛˌtinəˈsɔrəs/) was a prehistoric reptile genus belonging to the Pterosauria. It lived in the late Triassic period in the middle Norian (about 221-210 million years ago). Three fossils have been found near Cene, Italy. The first fossil, the holotype MCSNB 2886, is fragmentary and disarticulated. The second, the articulated paratype MCSNB 3359, lacks any diagnostic features of Peteinosaurus and thus might be a different species. This paratype has a long tail (20 cm) made more stiff by long extensions of the vertebrae; this feature is common among pterosaurs of the Triassic. The third exemplar is MCSNB 3496, another fragmentary skeleton. All specimina are those of subadults and of none has the skull been preserved. Like most pterosaurs, Peteinosaurus had bones that were strong but very light. Peteinosaurus is trimorphodontic, with three types of conical teeth. An insectivorous lifestyle has been attributed to Peteinosaurus. The fifth toe of Peteinosaurus was long and clawless. Its joint allowed it to flex in a different plane than the other phalanges, an adaptation whose function is unknown. Peteinosaurus is one of the oldest-known pterosaurs, and at a mere sixty centimetres, had a tiny wingspan when compared to some later genera, such as Pteranodon whose wingspan exceeded twenty feet. Its wings were also proportionally smaller than those of later pterosaurs, as its wing length was only twice the length of the hindlimb. All other known pterosaurs have wingspans at least three times the length of their hindlimbs. It also had single cusped teeth that lacked the specialized heterodonty present in the other Italian Triassic pterosaur genus, Eudimorphodon. All these factors converge to hint that Peteinosaurus belongs to a group that possibly represents the most basal known pterosaurs: the Dimorphodontidae, to which it was assigned in 1988 by Robert L. Carroll. The only other known member of that group is the later genus Dimorphodon, which lent its name to the family including both genera. Later cladistic analyses however, have not shown a close connection between the two forms. Nevertheless the possible basal position of Peteinosaurus has been affirmed by Fabio Marco Dalla Vecchia who suggested that Preondactylus, according to David Unwin the most basal pterosaur, might be a subjective junior synonym of Peteinosaurus. A 2010 cladistic analysis by Brian Andres and colleagues placed Peteinosaurus in Lonchognatha which includes Eudimorphodon and Austriadactylus as more basal.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, диморфодонтиды, птерозавры, рамфоринхоидеи |
| March 9th, 2012 | |
| 05:20 pm [industrialterro] [Link] |
Eudimorphodon Eudimorphodon was a pterosaur that was discovered in 1973 by Mario Pandolfi near Bergamo, Italy and described the same year by Rocco Zambelli. The nearly complete skeleton was retrieved from shale deposited during the Late Triassic (mid to late Norian stage), making Eudimorphodon the oldest pterosaur then known. It had a wingspan of about 100 centimetres (3.3 ft) and at the end of its long bony tail may have been a diamond-shaped flap like in the later Rhamphorhynchus. If so, the flap may have helped it steer while maneuvering in the air. Eudimorphodon is known from several skeletons, including juvenile specimens. Eudimorphodon showed a strong differentiation of the teeth, hence its name, which is derived from ancient Greek for "true dimorphic tooth". It also possessed a large number of these teeth, a total of 110 of them densely packed into a jaw only six centimeters long. The front of the jaw was filled with fangs, per side four in the upper jaw, two in the lower jaw, that rather abruptly gave way to a line of smaller multipointed teeth, 25 in the upper jaw, 26 in the lower jaw, most of which had five cusps, others three or even four. The morphology of the teeth are suggestive of a piscivorous diet, which has been confirmed by preserved stomach contents containing the remains of fish of the genus Parapholidophorus. Young Eudimorphodon had slightly differing dentition with fewer teeth and may have had a more insectivorous diet. The top and bottom teeth of Eudimorphodon came into direct contact with each other when the jaws were closed, especially at the back of the jaw. This degree of dental occlusion is the strongest known among pterosaurs. The teeth were multi-cusped, and tooth wear shows that Eudimorphodon was able to crush or chew its food to some degree. Wear along the sides of these teeth suggests that Eudimorphodon also fed on hard-shelled invertebrates. Despite its great age Eudimorphodon has few primitive characteristics making the taxon of little use in attempting to ascertain where pterosaurs fit in the reptile family tree. Basal traits though, are the retention of pterygoid teeth and the flexibility of the tail, which lacks the very long stiffening vertebral extensions other long-tailed pterosaurs possess. Paucity of early pterosaur remains has ensured that their evolutionary origins continues to be a persiseant mystery with some experts suggesting affinities to dinosaurs, archosauriformes, or prolacertiformes. Within the standard hypothesis that the Dinosauromorpha are the pterosaurs' close relatives within an overarching Ornithodira, Eudimorphodon is also unhelpful in establishing relationships within Pterosauria between early and later forms because then its multicusped teeth should be considered highly derived, compared to the simpler single-cusped teeth of Jurassic pterosaurs, and a strong indicator that Eudimorphodon is not closely related to the ancestor of later pterosaurs. Instead it is believed to be a member of a specialized off branch from the main "line" of pterosaur evolution, the Campylognathoididae. Eudimorphodon currently includes two species. The type species, E. ranzii, was first described by Zambelli in 1973. It is based on holotype MCSNB 2888. The specific name honours Professor Silvio Ranzi. A second species, E. rosenfeldi, was named by Dalla Vecchia in 1995 for two specimens found in Italy. However, further study by Dalla Vecchia found that these actually represented a distinct genus, which he named Carniadactylus in 2009. One additional species is still currently recognized: E. cromptonellus, described by Jenkins and colleagues in 2001. It is based on a juvenile specimen with a wing span of just 24 centimeters, MGUH VP 3393, found in the early nineties in Greenland. Its specific name honors Professor Alfred W. Crompton; the name is a diminutive because the exemplar is so small. In 1986 fossil jaw fragments containing multicusped teeth were found in Dockum Group rocks in western Texas. One fragment, apparently from a lower jaw, contained two teeth, each with five cusps. Another fragment, from an upper jaw, also contained several multi-cusped teeth. These finds are very similar to Eudimorphodon and may be attributable to this genus, although without better fossil remains it is impossible to be sure.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее )
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 05:06 pm [industrialterro] [Link] |
Caviramus Caviramus is a genus of "rhamphorhynchoid" pterosaur from the Late Triassic (late Norian-early Rhaetian-age) lower Kössen Formation of the Northern Calcareous Alps of Switzerland. The genus was in 2006 named by Nadia Fröbisch and Jörg Fröbisch. The type species is Caviramus schesaplanensis. The genus name is derived from Latin cavus, "hollow" and ramus, "branch". The specific name refers to Mount Schesaplana. The genus is based on holotype PIMUZ A/III 1225, three non-contiguous fragments of a ramus (lower jaw) of the mandible with multicuspate teeth. Two teeth are preserved, one with three cusps, and one with four; despite this difference the authors consider them as essentially isodont. The number of teeth is estimated at a minimum of twelve and a maximum of seventeen. A row of large oval foramina runs parallel to the tooth row; foramina in the form of small holes in the anterior part of the lower jaw suggest some sort of soft-tissue structure, or a keratin covering. The jaw is light and hollow. The teeth of this genus resemble those of Eudimorphodon, but the jaw is different. The discovery of this genus is a find of some significance, as there are few pterosaurs known from the Triassic. A second specimen, originally assigned to its own genus and species as Raeticodactylus filisurensis, consists of a single disarticulated partial skeleton including an almost complete skull. The skull shows that it had a tall thin bony crest running along the midline of the front of the upper jaw, and a keel on the lower jaw. The teeth at the front of the upper jaw, in the premaxillae, were fanglike, whereas the teeth in the upper cheeks (the maxillae) had three, four, or five cusps, similar to those of Eudimorphodon. Caviramus had a wingspan of about 135 centimeters (53 in), and may have been a piscivore, potentially a dip-feeder. Despite the resemblance to Eudimorphodon the authors classified Caviramus as Pterosauria incertae sedis. A 2009 study by Fabio Dalla Vecchia concluded that Raeticodactylus, which is known from a more complete skeleton including lower jaw, probably belong to the same genus, and possibly the same species, if the differences (such as size and the presence of a crest in the Raeticodactylus specimen) are not due to sex or age. Subsequent studies have supported their synonymy. Dalla Vecchia found the two forms in a sister clade of Carniadactylus, implying that Caviramus was a member of the Campylognathoididae.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 04:55 pm [industrialterro] [Link] |
Austriadactylus Austriadactylus is a genus of "rhamphorhynchoid" pterosaur. The fossil remains were unearthed in Late Triassic rocks of Austria. The genus was named in 2002 by Fabio Marco Dalla Vecchia e.a.. The type species is Austriadactylus cristatus. The genus name is derived from Latin Austria and Greek daktylos, "finger", in reference to the wing finger of pterosaurs. The specific epithet means "crested" in Latin, a reference to the skull crest. The genus is based on holotype SMNS 56342, a crushed partial skeleton on a slab, found in an abandoned mine near Ankerschlag in Tirol, in the Norian Seefelder Beds. The counterslab has been lost and with it some of the bone. The fossil consists of the skull, lower jaws, some vertebrae, parts of the limbs and pelvic girdle, and the first part of the tail. The elongated skull has a length of eleven centimetres. It carried a bony crest that widened as it descended towards the snout, up to height of two centimetres. The triangular nares formed the largest skull openings. The also triangular fenestrae antorbitales are smaller that the orbits. The teeth differ in shape and the species was thus heterodont. Most teeth are small and tricuspid or three-pointed. In the front of the upper jaw five larger recurved teeth with a single point form a prey grab; six or seven such teeth are also interspersed with the smaller teeth more to the back of the mouth. There are at least seventeen and perhaps as much as 25 tricuspid teeth in the upper jaw, for a total of perhaps 74 teeth of all sizes in the skull. The number of teeth in the lower jaws cannot be determined. The flexible tail did not have the stiffening rod-like vertebral extensions present in other basal pterosaurs. The wingspan has been estimated at about 120 centimetres. Austriadactylus was in 2002 assigned by the describers to a general Pterosauria incertae sedis, but some later analyses showed it to have been related to Campylognathoides and Eudimorphodon in the Campylognathoididae. It has even been suggested it was a junior synonym of Eudimorphodon, though perhaps a distinct species in that genus.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры, рамфоринхоидеи |
| 04:27 pm [industrialterro] [Link] |
Scleromochlus Scleromochlus is an extinct genus of small avemetatarsalian from the Late Triassic period. A lightly built cursorial animal, its phylogenetic position has been debated; as different analyses have found it to be either the basal-most ornithodiran, the sister-taxon to Pterosauria, or a basal member of Avemetatarsalia that lies outside of Ornithodira. Scleromochlus is a monotypic genus (single species), which is the type species S. taylori. Scleromochlus taylori was about 181 mm (about 7.1 inches) long, with long hind legs; it may have been capable of four-legged and two-legged locomotion. Its fossils have been found in the Carnian Lossiemouth Sandstone of Scotland. The holotype is BMNH R3556, a partial skeleton preserved as an impression in sandstone; part of the skull and tail are missing. Птероза́вры (Pterosauria — «летающие ящеры») — отряд вымерших летающих архозавров. Жили в мезозое на протяжении более 200 млн лет. Последние представители относятся ко времени вымирания динозавров, около 65 млн лет назад. В середине XVIII в. учёные извлекли из юрских зольнхофенских известняков (Бавария, Германия) несколько полных скелетов небольшого животного. Это существо настолько отличалось от всех современных животных, что долгие годы учёные терялись в догадках, что это за животное. Одни сочли его неведомой уткой, другие — летучей мышью и даже миниатюрным драконом. И только в 1809 году великий французский зоолог и палеонтолог Жорж Кювье окончательно установил, что найденные скелеты принадлежали вымершей летающей рептилии. Кювье назвал её птеродактилем («пальцекрыл»). Позднее, в XIX веке вокруг птерозавров развернулась ещё одна дискуссия. Ведущие британские учёные того времени Ричард Оуэн и Гарри Сили спорили о том, были ли птерозавры холоднокровными рептилиями или теплокровными «примитивными птицами». Картину прояснили последующие ископаемые находки. Без сомнения, птерозавры являлись рептилиями (как и полагал Оуэн), но теплокровными. Размах крыльев птерозавров достигал 7 метров, что более чем вдвое превышает этот показатель у любой из ныне живущих птиц. Их крылья представляли собой складки кожи, натянутые между боками тела и очень длинным четвёртым пальцем передних конечностей. Скелет обладал облегченной конструкцией: тонкие полые кости, лёгкий череп почти что сетчатой структуры, грудина имела киль, как у птиц. Строение скелета крыльев и плечевого пояса указывает на наличие мощной махательной мускулатуры. Необходимость в полёте возникла в связи с питанием рыбой и трупами, плавающими в воде, но птерозавры питались также и насекомыми. В настоящее время споры о способности птерозавров летать не утихают. Одни специалисты полагают, что гигантские рептилии могли преодолевать огромные расстояния по воздуху, а другие утверждают, что если и могли, то лишь в тихую погоду. На полуводный образ жизни указывают перепончатые лапы. Вытянутые в клюв челюсти могли нести зубы. У некоторых поздних форм, например, птеранодонов, были беззубые челюсти. Зубы некоторых видов свидетельствуют о питании планктоном. Птерозавры имели достаточно хорошо развитый мозг (преимущественно за счет мозжечка, отвечающего за координацию движений) и острое зрение. У них были редуцированны (уменьшены) обонятельные доли и развиты зрительные доли, что говорит о том, что летучих ящеров больше интересовало, как что-то выглядело, нежели как оно пахло. Тело было покрыто своеобразной «шерстью», что позволяет делать предположения о теплокровности птерозавров. На теплокровность указывает и обильное кровоснабжение костей, подобное тому, что наблюдается у птиц и млекопитающих. Возможно, что «шерсть» — это «пуховое оперение» молодых животных, так как у многих видов (например, Eudimorphodon) следы власоподобных структур обнаружены только на останках молодых особей. Птерозаврам для целей минимизации проблем с терморегуляцией при полноценном бодрствовании в светлое время суток просто необходимо было иметь многослойную, сложно устроенную мембрану крыла с развитой сосудистой сетью, обеспечивающей кондиционирование, а также наружные покровы, способные защитить от попадания избыточного количества прямых и отражённых солнечных лучей (например, во время полётов над водой), и гарантировать доставку крови к голове одной и той же температуры. По причине высокой специализации анатомии птерозавров для полета и отсутствия на данный момент известных переходных эволюционных форм, предки этих рептилий до конца не понятны. По некоторым гипотезам, они происходят от орнитодир, таких как Scleromochlus, относящихся к Avemetatarsalia близких к эупаркерии или же от проторозавров (особенно примечательным в этом предположении является вид шаровиптерикс).
Репродукции (1, 2, 3, 4, 5, 6):
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, птерозавры |



















/011_Gnathosaurus.jpg)























































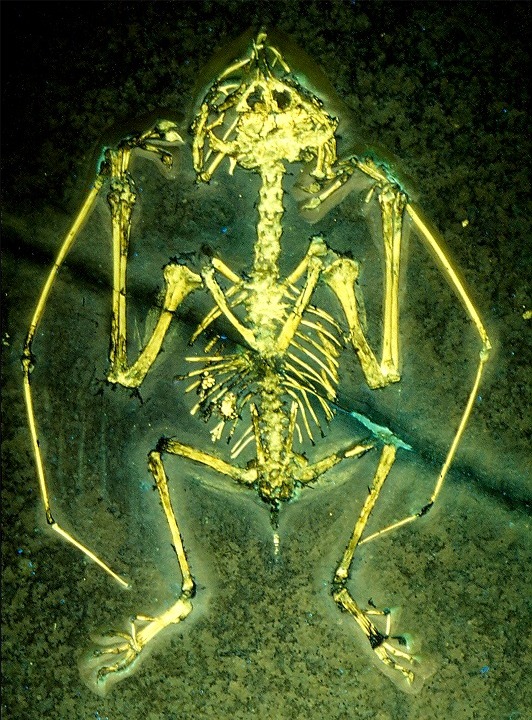

















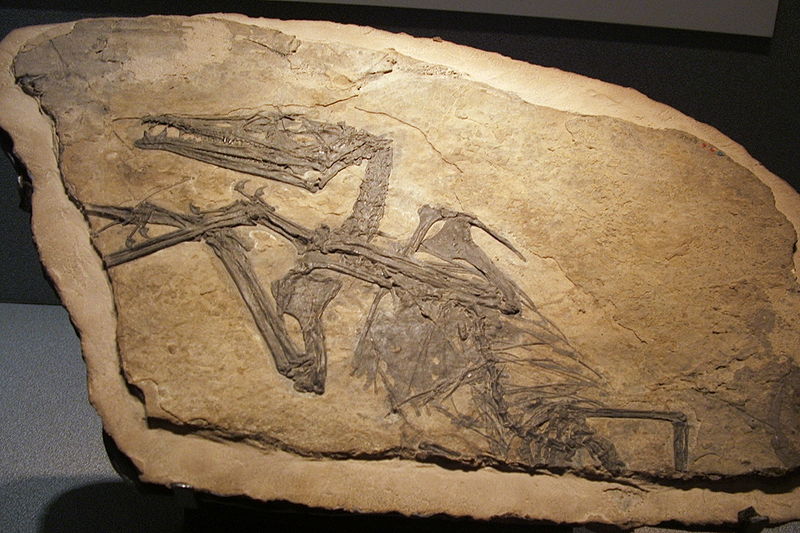
.JPG)














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}