[Recent Entries][Archive][Friends][User Info]
Below are the 20 most recent journal entries recorded in the "Сообщество, посвящённое ра" journal:| March 30th, 2012 | |
|---|---|
| 08:34 pm [industrialterro] [Link] |
Триасовое вымирание Триасово–юрское вымирание отмечает границу между триасовым и юрским периодами, 199,6 миллионов лет назад и является одним из крупнейших вымираний мезозойской эры, глубоко затронувших жизнь на Земле и в океанах. Целый класс конодонтов, составлявших 20% от всех морских семейств, все широко распространённые круротарзы (crurotarsans) (не динозавроподобные архозавры), некоторые из остававшихся терапсид и многие виды из широко распространённой группы земноводных полностью исчезли. По меньшей мере половина известных сейчас видов, живших на Земле в то время, вымерли. Это событие освободило экологические ниши, позволив динозаврам полностью доминировать на суше, начиная с юрского периода. Триасовое вымирание произошло менее чем за 10 000 лет и происходило непосредственно перед тем как Пангея начала распадаться на части. Статистический анализ потерь среди морской фауны в это время наводит на мысль, что уменьшение разнообразия было связано, скорее, со спадом в темпе видообразования, чем с ростом вымирания. Было предложено несколько объяснений этого события, но все они не в полной мере отвечают предъявленным требованиям: Постепенная перемена климата или флуктуации уровня океана в течение позднего триасового периода. Однако, это не объясняет внезапность вымирания существ в океане. Падение астероида, но нет датированного этим временем ударного кратера, образование которого совпадало бы с триасово–юрской границей (столкновение ответственное за кольцеобразную структуру Маникуаганского озера произошло за 12 миллионов лет до триасово–юрского вымирания). Массовые извержения вулканов, особенно излияние базальтовых лав в Центрально-атлантической магматической области (Central Atlantic Magmatic Province или CAMP), высвободило бы в атмосферу углекислый газ или диоксид серы которые в свою очередь явились бы причиной сильного глобального потепления (от первого газа) или похолодания (от второго газа). Гипотеза о метангидратном ружье. Потепление из-за вулканизма и накопления углекислого газа в атмосфере, привело к высвобождению метана из донных клатратов. Выделение метана, даже более сильного парникового газа чем CO2, ускорило потепление еще сильнее, что в свою очередь привело к ещё большему высвобождению метана со дна океанов. Этот процесс мог привести к быстрому изменению глобальной температуры. Изотропная структура окаменелых почв позднего триасового и раннего юрского периодов не демонстрируют доказательств, каких-либо изменений количества углекислого газа (СО2) в атмосфере. Однако позднее, некоторые доказательства все же были найдены в почвах относящихся к триасово–юрской границе, предполагая, что тогда произошло увеличение количества атмосферного СО2 и некоторые исследователи предположили, что причиной этого роста и, как следствие, массового вымирания могла послужить комбинация вулканического СО2, выделившегося из лав, и катастрофическое разложение газовых гидратов. Кроме того, газовые гидраты были предложены как одна из возможных причин массового пермского вымирания в конце пермского периода.
Tags: Массовое вымирание, Триас |
| 08:11 pm [industrialterro] [Link] |
Немного триасовой ботвы С началом верхней перми начался новый период эволюции растительного царства - мезофит. Примитивные сосудистые споровые растения вымирают и замещаются голосеменными, становящимися главенствующим элементом флоры. Наиболее широко распространенными представителями последних в начале мезофита были цикадовые и беннетиты. Некоторые представители цикадовых, иначе называемых саговыми, изображены в левой части картины. Стебли цикадовых были короткими и клубневидными или прямыми столбообразными, изредка разветвленными. На вершине они несли крону из крупных перистых листьев, напоминающих листья пальм. Беннетиты связаны с цикадовыми тесным родством. Среди них имелись как кустарники (Wielandiella), так и древесные растения (Williamsonid). Широко распространились в мезофите также гинкговые и хвойные - ели, кипарисы и сосны (изображены на заднем плане). Появляются гигантские мамонтовые деревья, или секвойи. На влажных местах продолжают существовать папоротники (задний план, справа), а на заболоченных участках - хвощи (средний план, справа), однако все эти криптогамные растения сильно мельчают.
Репродукции (1, 2):
Типичный пейзаж европейского нижнего триаса. В это время в Европе господствовали условия засушливого климата. Растительность повсеместно отступала к воде. По берегам небольших водоемов росла Pleuromeia, относящаяся к плаунам и достигающая высоты 2 м. Ее стройные стволы несут рубцы - следы опавших листьев и увенчаны густой лиственной кроной, на самой вершине шишковидный спорофил. В воде растет близкая к хвощам Schizoneura, на заднем плане изображена группа хвойных рода Voltzia. На прибрежном песке отпечатались следы неизвестного ящера, ранее относимого к роду Chirotherium. Сегодня ученые полагают, что эти следы принадлежат текодонту тицинозуху {Ticinosuchus). Текодонты были предками динозавров, летающих ящеров, крокодилов и птиц..
Триас Европы. Слева похожие на столбы Pleuromeiа, на заднем плане - примтивные хвойные рода Voltzia. Справа крупный плотоядный ящер из отряда текодонтов рода Ticinosuchus и его следа, напоминающие отпечатки человеческих рук.
Ландшафт, типичный для верхнего триаса - нижней юры Северного Полушария.
Tags: Растительность, Триас |
| March 29th, 2012 | |
| 09:59 pm [industrialterro] [Link] |
Megazostrodon Мегазостродон (Megazostrodon) — вымершее животное, найденное в Южной Африке. Единственный известный представитель семейства Megazostrodontidae. Признаётся одним из первых представителей млекопитающих, поскольку найдено в отложениях, чей возраст определён примерно в 200 миллионов лет. Имеет некоторые характеристики, не вполне присущие млекопитающим, которые однако не так уж и велики, что позволяет утверждать, что данный вид относится к последней стадии перехода от цинодонтов к настоящим млекопитающим. Мегазостродон был найден в позднем триасе Южной Африки. Он меньше морганукодона (Morganucodon), но такой же активный хищник, питавшийся мелкими наземными беспозвоночными. Образ жизни этого зверька напоминал современных землероек. Мегазостродон, одно из первых млекопитающих, имел лишь 10 см в длину. Первые млекопитающие появились примерно 220 миллионов лет назад. Это были триконодонты. К ним относится и мегазостродон, размером не больше мыши; он вёл ночной образ жизни, чтобы не попадаться хищникам. Питался насекомыми, червями, мелкими рептилиями и даже, вполне возможно, яйцами динозавров. Именно от таких мелких пушистых зверьков, подобных мегазостродону, произошли все современные млекопитающие. Как и его близкий родич морганукодон, мегазостродон был небольшим четвероногим созданием и, скорее всего, питался насекомыми и другими беспозвоночными, вроде червей. Скелет мегазостродона является самым известным среди скелетов ранних млекопитающих и демонстрирует образ тела с низким профилем, но прямой походкой. Достигая только 15 см в длину, этот крошечный зверёк был скорее всего ночным и теплокровным. Побочное доказательство теплокровия мегазостродона включает присуствие вторичного нёба. Палеонтолог Майкл Бентон считает, что разделяя дыхательное горло с глотательным, это вторичное нёбо позволяло животным быстро дышать при кормёжке. Этот метод поедания пищи можно увидеть у многих самых активных теплокровных животных современности. Во многом другом мегазостродон напоминал современных насекомоядных. Его черепная коробка, которая указывает на увеличение частей мозга ответственных за слух и обоняние, сильно намекает на его ночной образ жизни, т.к. и то, и другое чувство особенно полезно для ночных животных.
|
| 09:40 pm [industrialterro] [Link] |
Morganucodon Морганукодон (Morganucodon, «зубы из Гламоргана», по латинскому названию этой местности в «Книге Судного дня») — одно из древнейших млекопитающих, существовавшее в позднем триасе и предположительно вымершее в раннем юрском периоде. Впервые его останки обнаружил в Гламоргане Вальтер Георг Кюне.) Род обитал на Земле 221 - 210 млн лет назад. Отождествляется с эострозодоном. В отличие от многих других ранних млекопитающих, морганукодон хорошо представлен многочисленным и хорошо сохранившимся, хотя и в значительной мере расчленённым, материалом. Большая часть материала происходит из Гламоргана в Уэльсе (Morganucodon watsoni), но ископаемые останки также найдены в провинции Юньнань в КНР (Morganucodon oehleri) и во многих других местах в Европе и Северной Америке. Некоторые, по-видимому, близкородственные животные (мегазостродон) известны по уникальным останкам, найденным в Южной Африке. Как и большинство ранних млекопитающих, морганукодон был небольшим, покрытым шерстью животным, опиравшимся на всю стопу. Хвост был относительно длинным. Согласно Кемпу (Kemp, 2005) «длина черепа составляла 2-3 см, длина тела около 10 см. Внешне напоминал мышь или землеройку». Морганукодон, по всей видимости, вёл ночной образ жизни, днём скрываясь в норе. Рацион состоял, по-видимому, из насекомых и других мелких животных. Скорее всего, морганукодон откладывал яйца. Морганукодон (Morganucodon Kuhne, 1949) - типовой и самый известный род семейства Morganucodontidae, появившийся в палеонтологической летописи около 210 млн лет назад и наиболее хорошо изученное триасовое млекопитающее. Все разновидности охарактеризованы примитивными особенностями линейного выравнивания трёх основных бугорков, присутствия лингвального цингулида и связанных выступов, а также присутствием лингвальных и буккальных бугорков на поверхности зубов. Этими примитивными характерами Morganucodon может быть отделен от всех эутриконодонтов. Смежные нижние коренные зубы совмещаются размещением дистального острого выступа в вырезе между средними выступами последующего зуба, таким образом отличая Morganucodon от Dinnetherium, Kuehneotherium, и эутриконодонтов. У всех разновидностей Morganucodon присутствует характерный окклюзионный образец коренного зуба: центральный острый выступ нижнего моляра закрывает с лингвальной стороны впадину между выступами верхнего. Центральный выступ верхнего закрывает на буккальной стороне впадину между острыми выступами нижнего. Этими окклюзионными особенностями Morganucodon легко отличим от Megazostrodon, Brachyzostrodon, и Indozostrodon. Также наличествует много типичных нижнечелюстных особенностей основных млекопитающих: присутствие мобильного симфиза, присутствие постдентарной ложбинки, слабо развитая внешняя массетерная ямка. Морганукодон был зверьком 15-20 см длиной, весом около 20-30 гр с узкой мордой и удлинёнными челюстями, усаженными мелкими и острыми трехбугорчатыми зубами. Череп длиной 3 см, снабженный твердым нёбом. Голова по отношению к туловищу выглядит крупной, а конечности — тонкими. В общих чертах выглядел похожим на землеройку или мышь. Крошечные ямки на морде говорят о наличии вибрисс, крупные глазницы - о сумеречной или ночной активности. Уши имели сложное строение. Челюсти двигались не просто вверх и вниз, вперёд и назад, как у цинодонтов, а по более сложной треугольной траектории, будучи уже типичными зубами млекопитающего. Детские молочные зубы сменялись набором постоянных. Строение зубов показывает, что морганукодон был больше плотоядным, чем всеядным. Конфигурация суставных поверхностей нижних частей конечностей указывает, что постановка и характер движения последних напоминали свойственные примитивным ныне живущим слабокурсориальным млекопитающим типа сумчатого Didelphis и плацентарного Tupaia. Первый, или большой (hallux), палец задней конечности, возможно, несколько противопоставлялся остальным. Этот признак, а также наличие хорошо развитых когтей указывают на возможный древесный образ жизни. Однако данные признаки могли также возникнуть и в любой другой сложной для локомоции среде, заставляющей часто хвататься, цепляться и карабкаться, когда функциональные различия между наземным и древесным образом жизни невелики. Сумчатые кости не найдены, но поскольку они встречаются и у тритилодонтов, и нескольких других групп примитивных млекопитающих, то, возможно, были и у морганукодона.
|
| 09:12 pm [industrialterro] [Link] |
Eozostrodon Eozostrodon was one of the earliest mammals. It lived during the late Triassic and the early Jurassic, about 210 million years ago. Eozostrodon was one of the largest early mammals, measuring more than a meter long. As with most early mammals, the classification of Eozostrodon is uncertain. Like modern monotremes, this mammal laid eggs. Young were then fed with milk from their mother's mammary glands. Its teeth were typically mammalian, being differentiated into molars and premolars with triangular cusps. With its long snout, four legs, eight toes, clawed paws and a long hairy tail, Eozostrodon resembled a shrew. The Morganucodonts ("Glamorgan teeth") are an extinct group of important animals that seem to represent the earliest of mammals. These were shrew-sized, likely warm-blooded and possibly possessed mammary glands, though not organized into mammae with nipples. The morganucodonts were most likely insectivorous and nocturnal, seeking prey while avoiding predators that were usually active by day. Such a trait would be inherited by their descendants to avoid the predatory dinosaurs. They existed for a large portion of the Mesozoic era, from the late Triassic to early Cretaceous periods. Unlike Sinoconodon and the therapsids, their teeth are diphyodont (meaning that they possessed two sets of teeth as in all living mammals) and not polyphydont (meaning that the teeth are constantly replaced, as in reptiles). Furthermore, the primitive postcanine teeth found in more archaic proto-mammals have been replaced by true molars and premolars. The teeth are structured in such a way that a one-to-one occlusion and wear facets are present. The septomaxilla, a primitive feature also found in Sinoconodon, is present as well as a fully ossified orbitosphenoid. The anterior lamina is enlarged. The cranial moiety of the squamosal is a narrow bone that is superficially placed to the petrosal and parietal. Unlike its predecessors, the morganucodonts have a larger cerebral capacity and a longer cochlea. The atlas elements are unfused, there is a suture between the dens and axis. The cervical ribs are not fused to the centra. The coracoid and procoracoid, which are absent in therians, are present. The head of the humerus spherical as in mammals, but the spiral ulnar condyle is cynodont-like. In the pelvic girdle, the pubis, ilium and ischium are unfused. Scientists still debate if the morganucodonts should be classified as true mammals or classified as a clade outside of mammalia. An argument that is often used to classify them as non-mammals is the fact that they did not possess the three middle ear bones, they were equipped with a double jaw-joint instead (see above). Morganucodonts may be tagged as "prototherians."
|
| March 27th, 2012 | |
| 09:35 pm [industrialterro] [Link] |
Sinoconodon Синоконодон (Sinoconodon «китайский конический зуб») — вымершее животное, протомлекопитающее поздего триасового периода. Окаменелости найдены в Китае, провинция Юньнань, возраст — около 208 млн лет. Самым близким известным родственником синоконодона был, вероятно, морганукодон (возраст останков — 205 млн лет). Тем не менее, синоконодон существенно отличается от других маммалиаформов: его рост, по-видимому, не прекращался в течение всей жизни, точная окклюзия (смыкание зубов) отсутствует, что является примитивным признаком. Большинство зубов постоянно менялись, кроме передних щёчных, которые в случае утраты не восстанавливались и ни на одном известном черепе не сохранились. Синоконодон в целом считается более примитивным, обладает менее выраженными признаками млекопитающих, чем родственные ему маммалиаформы, включая морганоконудонтов и докодонтов, которые появились через несколько миллионов лет, и, вероятно, является реликтом более ранней стадии эволюции млекопитающих. В связи с этим, в одних работах он упоминается как самое примитивное млекопитающее, в других вовсе не считается млекопитающим. Синоконодон (Sinoconodon Patterson и Olson, 1961) – позднетриасовое млекопитающее примерно 15 см длиной. Вероятно, имел маленькое, пушистое, подобное полёвке тело с относительно крупной головой (приблизительно 60 мм в длину и 23 мм в ширину) с большими глазами и рептильными зубами, постоянно сменяющимися на протяжении всей жизни животного. Sinoconodon обладал примитивными "триконодонтными” щечными зубами, приспособленными к хищническому или насекомоядному образу жизни. К сожалению, постчерепные останки не позволяют определить локомотивные особенности передвижения. Sinoconodon имел широкий онтогенетический диапазон изменения размеров черепа: наименьший известный череп приблизительно 20 мм длиной, а наибольший - крупнее, чем 60 мм. Постчерепные окаменелости Sinoconodon демонстрируют, что у него имелись развитые передние конечности и плечевой пояс, мало чем отличающиеся от таковых у цинодонтов (Jenkins, 1971; Kemp, 1982; Sues, 1983), и современных монотремат. Sinoconodon однозначно помещен в Mammalia, потому что разделяет много особенностей с другими млекопитающими - например, краниомандибулярный сустав, сформированный дентарным мыщелком и гленоидом сквамозной кости, мысок височной кости, её антериорная стенка и костистая впадина для узла тройничного нерва. Однако расположение зубов Sinoconodon примитивно. Sinoconodontidae сохранили примитивное чередование и многократные замены резцов и клыков, замеченных у многих цинодонтов. Со временем, антериорные постклыковые зубы последовательно выпадали, приводя к постепенно увеличивающейся постклыковой диастеме, идентичной таковой у гомфодонтных цинодонтид (Hopson, 1971). Нижнечелюстные структуры и структуры среднего уха - по существу те же самые, как у Morganucodon и Megazostrodon, с примитивной постдентарной ложбинкой (хотя более уменьшенной, чем у Morganucodon).
Crompton и Sun (1985) наглядно показали, что верхние и нижние постклыковые зубы Sinoconodon отличаются по размеру и что они имеют переменное число. Верхние и нижние постклыковые зубы не совмещаются, так как на их выступах нет никаких зон изнашивания. Это - примитивная особенность, которая принудила Crompton и Sun (1985) предположить, что Sinoconodon более примитивен, чем многие из родственных ему млекопитающих: Morganucodon, Megazostrodon, Kuehneotherium, докодонты и харамивиды. Основываясь на дополнительных характерах зубной замены, краниомандибулярного сустава и ушной области, Crompton и Luo (1993) предложили, что Sinoconodon должен являться сестринским таксоном по отношению к другим группам млекопитающих. Таким образом, Sinoconodontidae представляют одно из ранних боковых происхождений позднетриасовых-раннеюрских млекопитающих.
Млекопитающие имеют два апоморфия слухового региона: удлиненный улитковый канал и возвышенность на тимпанной стороне кохлеарной области, именуемого как мысок верхнего края пирамиды височной кости. У цинодонтов улитка короче, меньше и сориентирована более медиально, чем улитковый канал млекопитающих; их кохлеарное размещение сформировано предушной областью и базисфеноидом. Ископаемые материалы Adelobasileus и Sinoconodon подтверждают развитие у ранних млекопитающих кохлеарной области. Sinoconodon имеет полностью сформированный мысок верхнего края височной кости, но короткую улитку, которая расширена только на одну треть длины мыска. Кохлеарный канал Morganucodon – вдвое длиннее, чем у Sinoconodon как относительно черепа, так и мыска. Более поздние млекопитающие - как, например, Haldanodon и мультитуберкуляты, демонстрируют более длинный улитковый канал. Предполагается, что мысок, преобразованный расширением височной кости за счет базисфеноида (вероятно, базиокципитально) является свидетельством перехода цинодонтов в млекопитающие (Luo, Z.-X., A.W. Crompton, и S.G. Lucas, 1995). Это изменение в окостенении и ориентации кохлеарной области эффективно для лучшей акустической изоляции внутреннего уха.
Зубная формула: 5.1.2.5/5.1.2.5. Все типы зубов являются вариабельными и нумеруются вследствие цинодонтоподобных непрерывных зубных замен в корреляции с ростом черепа. Sinoconodontidae сохранили переменные замены резцов и клыков, замеченные у большинства цинодонтов. Клыки заменялись, по крайней мере, четыре раза. Размер функциональных клыков является очень переменным среди различных черепных останков (Crompton и Luo, 1993). В самой полной форме клык подобен кинжалу. Верхний клык является настолько большим, что достигает нижней границы нижней челюсти, но на ранней стадии роста коронка клыка может быть короткой и конической.
Фактическое число функционирующих резцов является переменным. У некоторых черепных останков наличествовало только три функциональных резца в результате продолжающихся замен. Резцы имеют большие и небольшие размеры. Переменное расположение маленьких и больших зубов типично для цинодонтов, имевших непрерывную и переменную замену. Премоляры только присутствуют в ювенильных экземплярах (Zhang и др., 1998). Первый премоляр, кажется, потерян без замены у старших экземпляров. Второй премоляр имеет листообразную форму у молодых особей, и впоследствии исчезает у более старших возрастов. Нижние постклыковые зубы вообще "триконодонтоподобны", но демонстрируют более широкий диапазон морфологического изменения, возможно связанного с их заменой. У предпоследних моляров некоторых больших черепов имеется срединный цингулидный бугорок, который отсутствует у моляриморфного зуба меньших черепов. Этот выступ является переменной особенностью и может отсутствовать в антериорных постклыковых зубах. У верхнего моляра меньших экземпляров есть "триконодонтоподобная" трёхвершинная коронка с острыми выступами. В отличие от этого, верхний моляр больших экземпляров имеет тенденцию развивать дополнительный дистальный бугор. Как упомянуто ранее, у Sinoconodon вследствие вариаций в зубных размерах и морфологии между верхними и нижними постклыковыми зубами окклюзии нет.
Sinoconodon обладает многими характерами, предполагающими, что это - основная форма, относящаяся к Mammalia. Дальнейшие экземпляры (Crompton и Luo, 1993) в достаточной мере подтвердили это заключение. Подтверждение его статуса обеспечено дентальным мыщелком и гленоидной ямкой сквамозной кости, полнотой постгленоидного гребня и расширением черепа. В качестве мозаичных характеристик можно указать диапазон примитивных характеров, имеющихся у тритилодонтид и трителодонтов. Самым важным является расположение зубов - сравнение ряда экземпляров отличающихся размеров показывает, что способ замены зубов был аналогичен цинодонтам, а не млекопитающим (Zhang и др. 1998; Kielan-Jaworowska и др. 2004). Резцы и клыки сменялись несколько раз, в то время как антериорные премоляры выпадали, но не заменялись.
Филогенетично Sinoconodon был, несомненно, близок к основной, относящейся к млекопитающим радиативной линии, сохранив примитивные характеры. Действительно, не принимая во внимание малоизвестного Adelobasileus и харамивид, он универсально принят как сестринская группа остальной части Mammalia. Как и у морганукодонтов, между орбитами Sinoconodon есть костистое разделение и сустав между сквамозной и зубной костями, но они более уменьшены, чем у морганукодонтид (Crompton и Sun, 1985). Однако, Sinoconodon, кажется, более примитивен, чем Morganucodon. Кохлеарная структура была более примитивной (Luo и др., 1995), и отсутствие последовательных зон изнашивания указывает, что Sinoconodon испытывал недостаток в смыкании коренных зубов. Это напоминает цинодонтов в сохранении большой септомаксиллярной кости и многократной замене резцов и клыков. Данные особенности предполагают, что Sinoconodon отклонился от относящегося к млекопитающим эволюционного ствола ранее, чем морганукодонтиды.
 Tags: Вымершие млекопитающие, Триас |
| 09:18 pm [industrialterro] [Link] |
Adelobasileus Аделобазилевс (Adelobasileus, от др.-греч. ἄδηλος βασιλεύς «неясный царь») — вымершее протомлекопитающее позднего триасового периода (ок. 225 млн лет назад). Хищник, размером приблизительно с мышь. Известен по одному черепу, найденному на территории Техаса в 1989 году палеонтологами из Музея естественной истории и науки Нью-Мексико, где и хранится находка. Особенности строения черепа, особенно устройство внутреннего уха, свидетельствуют о том, что аделобазилевс был переходной формой между цинодонтами и триасовыми млекопитающими. Аделобазилевс жил приблизительно на 10 млн лет раньше наиболее продвинутых в эволюционном плане цинодонтов и всех других известных маммалиаформов (Mammaliaformes). Это даёт основания предполагать, что он мог быть общим предком млекопитающих, или, что более вероятно, состоял в близком родстве с общим предком млекопитающих.
Tags: Вымершие млекопитающие, Триас |
| 07:41 pm [industrialterro] [Link] |
Protoavis Протоавис (Protoavis) — название рода, предложенное для окаменелостей позднего триаса (225—210 млн лет), обнаруженных на северо-западе Техаса (США). В 1991 году протоавис был описан палеонтологом Шанкром Чаттерджи (Sankar Chatterjee) как примитивная птица, однако эта интерпретация была подвергнута сомнению. Если интерпретация Чаттерджи верна, то протоавис является древнейшей из известных птиц, на 60-75 млн лет старше археоптерикса. Выделяется единственный вид Protoavis texensis («первоптица штата Техас»). Статус протоависа спорен. Некоторые палеонтологи (Е. Курочкин, Д. Петерз, Л. Мартин, Л. Хо) принимают птичий статус протоависа. Однако большинство палеонтологов отвергает связь протоависа с птицами и подвергает сомнению законность таксона. Согласно реконструкции Чаттерджи длина протоависа составляла около 35 см. Хотя он жил за 60-75 млн лет до археоптерикса, строение его скелета ближе к современным птицам. Из-за наличия зубов на клюве в нём предполагают хищника, размещение глаз на передней части черепа свидетельствует о ночном или сумеречном образе жизни. По мнению Чаттерджи, Протоавис был адаптирован к полету, на его костях присутствуют особые отметины — точки прикрепления перьев. Однако, по мнению других палеонтологов, небольшое количество окаменелостей не позволяет определить степень способности протоависа к полёту, более того, детальный их анализ не даёт никаких свидетельств наличия у него перьев. Е. Курочкин выделяет следующие особенности, связывающие протоависа с птицами.
Окаменелости протоависа найдены в 1984 в отложениях триасовой речной дельты. Они были смешаны с костями различных динозавров, что является возможным следствием массовой гибели при наводнении. Обнаруживший их учёный Шанкр Чаттерджи убеждён, что найденные им останки принадлежат двум особям одного вида - молодой и старой. К ним относятся лишь череп и фрагменты конечностей, достаточно диспропорциональные между собой, вследствие чего и появилась версия о химерности экземпляров, иными словами, гипотеза о том, что реконструированные скелеты, на самом деле, представляют собой скопление костей нескольких животных, принадлежащих к разным видам. Из-за небольшого числа находок и обстоятельств их обнаружения среди палеонтологов сохраняются сомнения относительно того, является ли протоавис птицей, и даже того, является ли он реально существовавшим животным. Российский палеорнитолог Е. Н. Курочкин, принимавший участие в изучении останков, полагает, что причина критического отношения учёных к протоавису носит преимущественно психологический характер, связанный с категоричностью утверждений Чаттерджи, который настаивает на способности обнаруженного существа к полёту. Принятие птичьего статуса протоависа подразумевает, что становление класса птиц произошло не позднее конца триасового периода. Следовательно, предков птиц следует искать среди триасовых архозавров. Это не согласуется с наиболее распространённой точкой зрения на филогению ранних птиц, согласно которой самыми близкими родственниками птиц являются троодонтиды и дромеозавриды — два семейства хищных динозавров (тероподов). Если данная филогенетическая теория верна, то эти семейства должны были возникнуть ещё в триасовом периоде, однако наиболее ранние окаменелости представителей этих двух семейств относятся к раннему мелу. По мнению Чаттерджи, ближайшими родственниками птиц являются триасовые цератозавры — группа тероподных динозавров, обладавшая рядом существенных особенностей, характерных для птиц (близкое к гетероцильному строение шейных позвонков, слияние тазовых костей и др.). Эти особенности скелета цератозавров традиционно расцениваются как результат конвергенции. Гипотеза дифилетического происхождения птиц, разрабатываемая Е. Н. Курочкином, частично основывается на изучении протоависа. Согласно этой гипотезе, веерохвостые птицы произошли от некоторой группы юрских архозавров, близким родственником которых является протоавис. Археоптерикс и энанциорнисы, согласно данной гипотезе, в свою очередь, являются потомками тероподов. Признание протоависа наиболее ранней на сегодняшний день птицей порождает временной промежуток в 85 млн лет между протоависом (210 млн лет) и первыми бесспорными веерохвостыми птицами (125 млн лет). В течение этих 85 млн лет (поздний триас, юрский период, ранний мел) должны были существовать примитивные птицы, окаменелостей которых однако не найдено. Исключение составляет археоптерикс (поздний юрский период), но он менее продвинут по сравнению с предполагаемой анатомией протоависа. Л. Уитмер заключил, что череп протоависа на самом деле принадлежит целурозавру. Эта оценка предполагает, что целурозавры появились не позднее конца триаса. Американские палеонтологи во главе с С. Чаттерджи (S. Chatterjee) при раскопках в районе Поста (штат Техас) обнаружили останки двух ископаемых позвоночных возрастом около 225 млн лет. Изучив находку, исследователи пришли к выводу, что имеют дело с наиболее древней среди известных науке птиц, жившей за 75 млн лет до археоптерикса, который, как известно, долгое время считался древнейшей первоптицей. Фрагментарность останков затрудняет определение и детальное описание находки, получившей название Protoavis. Ясно, однако, что эта предптица, как и археоптерикс, обладала когтистыми пальцами, хвостом и зубами, а следовательно, была близка к динозаврам. С другой стороны, в скелете протоависа найдена «вилочка» — грудная кость, встречающаяся только у настоящих птиц и отсутствовавшая у археоптерикса. В связи с этим Чаттерджи полагает, что протоавис обладал большей способностью к полету, чем археоптерикс; подкрепляется это предположение и строением костей крыла протоависа. Его череп весьма сходен с черепом современных птиц, а в задней части челюстей зубы отсутствуют. Возможно, протоавис стоял в начале эволюционной линии, приведшей древних птиц к постепенному избавлению от тяжелой, несущей зубы, челюсти и к способности свободного полета. Это предположение, однако, не полностью разделяет крупнейший палеонтолог и специалист по проблематике происхождения птиц Дж. Остром (J. Ostrom; Йельский университет, Нью-Хейвен, США), полагая, что имеющихся данных недостаточно, чтобы судить о столь значительной эволюционной «продвинутости» протоависа. С другой стороны, многие палеонтологи и раньше обращали внимание на то, что археоптерикс слишком примитивен и вместе с тем специализирован, чтобы служить прямым предком птиц. Кроме того, для этого ему «отпущено» слишком мало времени: ведь всего несколько миллионов лет спустя уже существовали настоящие птицы. Рядом с техасской находкой отпечатков оперения не обнаружено, однако на локтевых костях заметны выступы, к которым могли бы прикрепляться перья. Останки ископаемых птиц — вообще большая редкость, так как их хрупкие кости обычно плохо сохраняются. В данном случае обе найденные особи, вероятно, погибли при паводке или наводнении и их останки оказались захоронены под слоем ила. Chatterjee was convinced that some of these crushed bones belonged to two individuals – one old, one young – of the same species. However, only a few parts were found, primarily a skull and some limb bones which moreover do not well agree in their proportions respective to each other, and this has led many to believe that the Protoavis fossil is chimaeric, made up of more than one organism: the pieces of skull appear like those of a coelurosaur, while most parts of the limb skeleton suggest affinities to ceratosaurs and at least some vertebrae are most similar to those of Megalancosaurus, an avicephalan diapsid. If it really is a single animal and not a chimera, Protoavis would raise interesting questions about when birds began to diverge from the dinosaurs, but until better evidence is produced, the animal's status currently remains uncertain. Furthermore, palaeobiogeography suggests that true birds did not colonize the Americas until the Cretaceous; the most primitive lineages of unequivocal birds found to date are all Eurasian. Certainly, the fossils are most parsimoniously attributed to primitive dinosaurian and other reptiles as outlined above. However, coelurosaurs and ceratosaurs are in any case not too distantly related to the ancestors of birds and in some aspects of the skeleton not unlike them, explaining how their fossils could be mistaken as avian. Palaeontologist Zhonghe Zhou stated: "[Protoavis] has neither been widely accepted nor seriously considered as a Triassic bird ... [Witmer], who has examined the material and is one of the few workers to have seriously considered Chatterjee’s proposal, argued that the avian status of P. texensis is probably not as clear as generally portrayed by Chatterjee, and further recommended minimization of the role that Protoavis plays in the discussion of avian ancestry." Welman has argued that the quadrate of Protoavis displays synapomorphies of Theropoda. Paul has demonstrated the drepanosaur affinities of the cervical vertebrae. Gauthier & Rowe, and Dingus & Rowe have argued convincingly for identifying the hind limb of Protoavis as belonging to a ceratosaur. Feduccia has argued that Protoavis represents an arboreal "thecodont". In a study of early ornithischian dinosaurs, Sterling Nesbitt and others determined some of the partial remains of Protoavis to be a non-tetanuran theropod. The entire skull and neck are considered to be most likely from a drepanosaurid because the skull and neck are too big compared to the dorsal vertebrate of Protoavis.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| 06:48 pm [industrialterro] [Link] |
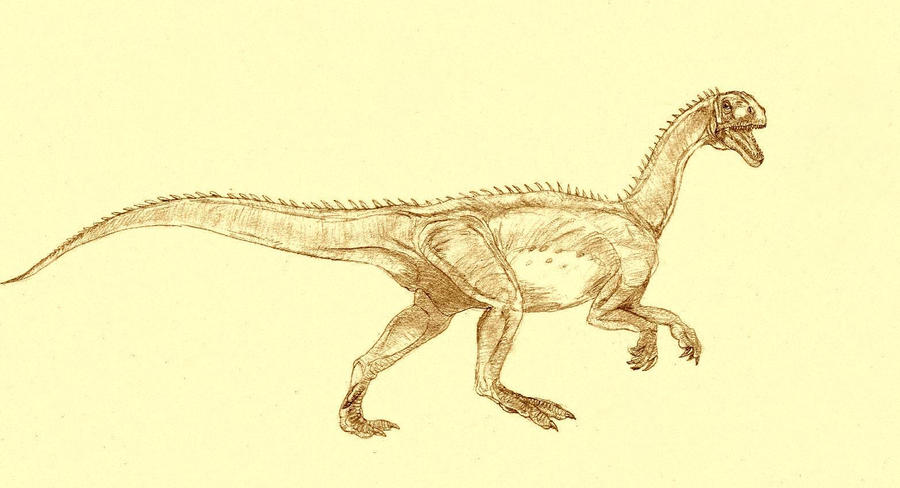
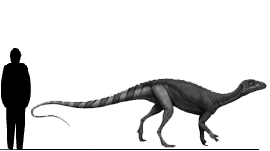
Zupaysaurus Зупайзавр (Zupaysaurus) — род вымерших ящеротазовых динозавров из подотряда тероподов, живших в позднем триасовом и раннем юрском периодах (около 216,5—203,6 миллионов лет назад), на территории нынешней Южной Америки. Окаменелости теропода были найдены в провинции Ла-Риоха (формация Los Colorados) в Аргентине. Впервые описан палеонтологами Аркуччи (Arcucci) и Кориа в 2003 году. Представлен одним видом — Z. rougieri. Хотя полный скелет до сих пор не был обнаружен, ученые считают Zupaysaurus двуногим хищником, достигавшим до 4 метров в длину. Также ученые предполагают возможное наличие двух параллельных гребней на его длиной морде. Zupaysaurus был динозавром среднего размера. Череп взрослой особи достигал 45 сантиметров в длину, что дает возможность ученым предположить общую длину животного, которая оценивается примерно в 4 метра от морды до кончика хвоста. Как и все известные тероподы, Zupaysaurus вероятно ходил только на задних конечностях, оставляя передние свободными для хватания добычи. Имел небольшой зазор, отделявший зубы предверхнечелюстных и верхнечелюстных костей верхней челюсти, таранная кость и кости пяточной лодыжки были слиты воедино, что отмечается также и у ранних тероподов. В верхней части черепа вероятно имел два тонких параллельных гребня, подобно другим тероподам, таким как Dilophosaurus и Megapnosaurus (Syntarsus) kayentakatae. Эти гребни предположительно были сформированы исключительно носовыми костями, в отличие от гребней многих других тероподов, у которых подобные образования дополнялись также и слезными костями. Ученые предполагают, что гребни, широко распространенные на черепах тероподов, возможно использовались ими в коммуникативных целях, таких как гендерное, групповое или видовое распознавание. Однако, более свежий анализ черепа подверг сомнению вероятность присутствия этих гребней в Zupaysaurus. Неопубликованное резюме, представленное на конференции, указывало на то, что структуры, первоначально идентифицированные как гребни, на самом деле, были фактически слезными костями, смещенными вверх во время процесса окаменения.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| 06:17 pm [industrialterro] [Link] |
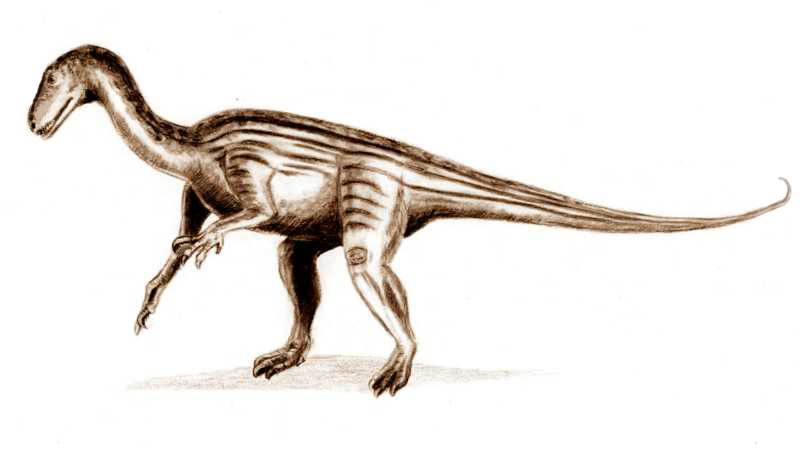
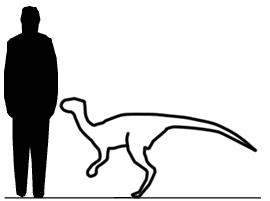
Unaysaurus Unaysaurus — вымерший род динозавров принадлежащих к группе Sauropodomorpha, живших в позднем триасе около 225 миллионов назад (карнийский—норийский ярусы) на юге нынешней Бразилии. Типовой вид - Unaysaurus tolentinoi. Ископаемые останки, голотип UFSM 11069, были найдены в 1998 году в окрестностях города Санта-Мария. Находка была сделана Толентино Марафигой случайно, во время прогулки (видовое название дано в его честь). Окаменелости залегали в слоях формации Caturrita, относящейся к карнийскому—норийскому ярусам, и они состоят из почти полного черепа и некоторых фрагментов туловища и конечностей. В 2004 году вид был описан в научном журнале Zootaxa Александром Кельнером. Название рода происходит от слова unay - «черная вода», на языке тупи, ссылаясь на регион Agua Negra, на территории которого была сделана находка. Unaysaurus был относительно небольшим травоядным динозавром, достигавшим в длину 2,5 метров и весившим 75 кг. Внешне напоминал прозауроподов: он передвигался на двух ногах (также не исключено, что он мог опираться и на 4 конечности) и имел длинную шею. В современных классификациях динозавра относят к семейству Plateosauridae, но вляется ли он вдействительности членом группы Prosauropoda в настоящее время остается неясным. Бразильские ученые обнаружили хорошо сохранившиеся окаменелые останки динозавра неизвестного ранее вида, который обитал на территории южной Бразилии 225 миллионов лет назад. Как передает Reuters, высота этого травоядного ящера, который передвигался на задних лапах, составляла около 70 сантиметров, а длина - 2,5 метра.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, платеозавриды, прозавроподы, ящеротазовые |
| March 26th, 2012 | |
| 08:47 pm [industrialterro] [Link] |
Thecodontosaurus Текодонтозавр (Thecodontosaurus — «ящер с собранными зубами») — род вымерших динозавров подотряда Sauropodomorpha. Назван из-за зубов, сидящих в очень отличительных гнездах. Очень примитивен. Внешне схож с анхизавром, за исключением более длинной шеи и хвоста и некоторых других особенностей. Этот род известен по более ста животным — как скелетам, так и отдельным костям, принадлежащих как молодым, так и взрослым животным длиной до 2,6 м и весом 40—70 кг. Обитал на территории нынешней Англии (поздний триас — ранняя юра, 215—195 млн.л.н.). From the fragmentary remains of Thecodontosaurus most of the skeleton can be reconstructed, the main exception being the front of the skull. Thecodontosaurus had a rather short neck supporting a fairly large skull with large eyes. Its jaws contained many small- to medium-sized, serrated, leaf-shaped teeth. This dinosaur's hands and feet each had five digits, and the hands were long and rather narrow with an extended claw on each. This dinosaur's front limbs were much shorter than the legs, and its tail was much longer than the head, neck and body put together. On average, it was 1.2 metres (3.9 ft) long, 30 centimetres (12 in) tall, and weighed 11 kilograms (24 lb). The largest individuals had an estimated length of 2.5 metres (8.2 ft). Michael Benton in 2000 noted the existence of a robust morph in the population, seen by him as a possible second species or, more likely, an instance of sexual dimorphism. Benton also indicated some unique derived traits, or autapomorphies for the species: a long basipterygoid process on the braincase; a dentary that is short in relation to the total length of the lower jaw; an ilium that has a back end that is subquadrate instead of rounded. The small size has been explained as an instance of insular dwarfism. Riley and Stutchbury originally saw Thecondontosaurus as a member of the Squamata, the group containing lizards and snakes. This did not change when Richard Owen in 1842 coined the Dinosauria, because Owen did not recognise Thecodontosaurus as a dinosaur; in 1865 he assigned it to the Thecodontia. Only in 1870 was Thomas Huxley the first to understand it was a dinosaur, though referring it incorrectly to the Scelidosauridae. Later it would normally be placed in either the Anchisauridae or its own Thecodontosauridae. Modern exact cladistic analyses have not been conclusive. Although not actually the earliest member of the group, Thecodontosaurus is sometimes placed in a very basal position among the sauropodomorph dinosaurs. It was earlier included under the Prosauropoda but more recently it has been suggested that Thecodontosaurus and its relatives were prior to the prosauropod-sauropod split. New reconstructions show that its neck is proportionally shorter than in more advanced early sauropodomorphs.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, ящеротазовые |
| 08:15 pm [industrialterro] [Link] |
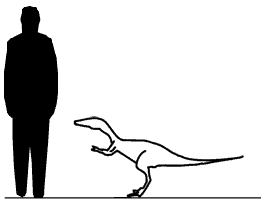
Tawa Тава (Tawa; в честь бога солнца народа пуэбло) — род динозавров подотряда тероподов позднего триасового периода. Обитал приблизительно 215—213 миллионов лет назад. Эти динозавры являются самыми ранними из тероподов на территории современной Северной Америки. Впервые обнаружен (частичный скелет) в 2004 году в карьере Хайден (Hayden Quarry) в Гост-Ранч (Ghost Ranch) на территории штата Нью-Мексико, а в 2006 году начались полномасштабные раскопки, в результате которых был найден почти полный ископаемый скелет Tawa, а также семь других скелетов, расположенных близко друг к другу. Вид Tawa hallae описан в декабре 2009 году Несбиттом (Nesbitt) и другими учеными. В их распоряжении оказался практически полный скелет молодой особи (голотип каталогизирован как GR 241), длиной около 2 метров. Изучение другого образца (каталогизированого как GR 242), также близится к завершению. Одна отдельная найденная бедренная кость позволяет предположить, что взрослые особи этого вида были, по крайней мере, на 30 % больше, чем исследованные молодые особи, в том числе и та из них, что является голотипом. При жизни динозавры данного вида были около 70 сантиметров в высоту. Взрослые особи, по расчетам исследователей, должны были достигать 4 метров в длину. Отличительной особенностью данного вида является очень длинный хвост. Исследователи подчеркивают, что им очень повезло. Дело в том, что останки не только прекрасно сохранились, но ещё и во время окаменения не были сдавлены породами. Таким образом, у исследователей имеется редкая возможность взглянуть на строение внутренних органов древних ящеров. Этот динозавр является одним из самых ранних тероподов на территории южной части нынешней Северной Америки. Изучение останков T. hallae по мнению ученых, поможет им прояснить эволюцию первых динозавров. Считается, что примерно 230 миллионов лет назад, когда все континенты были объединены в суперконтинент Пангею, древние ящеры впервые стали заселять разные регионы планеты. Новый вид динозавров помог определить, что тероподы возникли на территории Гондваны (региона, который позже стал Южной Америкой) и позже расселились по суперконтиненту. Также T. hallae помог определиться с классификацией другого динозавра — герреразавра. Набор некоторых характеристик у T. hallae окончательно устанавливают прямую связь между тероподами и герреразавром. Таким образом тава является промежуточным звеном между герреразаврами и неотераподами.
Размеры тела в сравнении с человеком (показан мелкий голотип и предполагаемый размер взрослой особи):
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| March 24th, 2012 | |
| 03:06 pm [industrialterro] [Link] |
Staurikosaurus Ставрикозавр (Staurikosaurus) — монотипный род тероподов позднего триасового периода из семейства Herrerasauridae, один из первых динозавров планеты. Единственный вид в роде - Staurikosaurus pricei. Родовое имя динозавра "ставрикозавр" связано с названием созвездия Южный Крест, изображённого на гербе Бразилии и видимого, наблюдаемого только в Южном полушарии, как свидетельство редких находок динозавров в Южном полушарии. Видовое имя pricei посвящено бразильскому палеонтологу Л.И.Прайсу (Llewellyn Ivor Price), обнаружившему скелет ставрикозавра. Ставрикозавр обитал в карнийском веке верхнего триасового периода, около 225 млн. лет назад. Он достигал высоты 2,25 м и длины 80 см и весил около 30 кг. Единственная находка ставрикозавра очень не полная: состоит из большей части позвоночника, фрагментов ног и нижней челюсти. Реставрация ставрикозавра выполнялась исходя из предположения, что динозавр на заре эпохи не мог быть очень сложным и иметь узко-специализированные органы. Поэтому ставрикозавра изображают обычно с простыми 5-ю пальцами передних и задних конечностей. Однако найденных фрагментов достаточно, чтобы судить о том, что ставрикозавр был довольно быстрым хищником. Ставрикозавр достигал длины 2,25 м и высоты 80 см и весил около 30 кг. Единственный экземпляр ставрикозавра был обнаружен при раскопках в районе Рио-Гранде-до-Сул в Южной Бразилии. Эдвин X. Кольберт из Американского музея естественной истории дал научное описание виду в 1970 году. Этот мелкий древний динозавр жил в позднем триасе, примерно 225 миллионов лет назад. До середины 20 века открытием останков динозавров изобиловало только Северное полушарие Земли, в то время как в Южном полушарии находок скелетов древних ящеров практически не было. Поэтому название динозавра происходит от созвездия Южного креста - в ознаменование находки динозавра в Южном полушарии, ведь до этого подобные находки были весьма редким явлением. Поэтому ставрикозавр - ящер "Южного креста". Ставрикозавр мог достигать примерно 2 метров в длину, что позволяет считать его достаточно мелким в сравнении с другими, более поздними динозаврами, и весил около 30-35 кг – это был очень легкий ящер. Зубы были заостренными, так что ставрикозавр скорее всего являлся хищником. Челюсть ящера имела интересное строение: она могла слегка выдвигаться вперед а также задвигаться глубже в пасть. Вероятно, таким образом, ставрикозавру было легче удерживать попавшую ему в пасть добычу. Относительно небольшое тело ставрикозавра венчала большая голова; хвост был тонким и длинным. Скорее всего, он служил своеобразным рулём, помогая ставрикозавру во время бега. Передвигался этот динозавр очень быстро. Палеонтологи не без оснований считают, что ставрикозавр мог быть самым быстрым ящером своего времени: на это указывают его длинные и сильные задние конечности. Немногое известно об этом мелком животном, но это очень важная находка, так как до самого последнего времени ставрикозавр считался самым древним из известных динозавров. Судя по его внешнему виду, он больше походил на мелких хищных теропод, но некоторые черты строения скелета роднили его с растительноядными прозауроподами. Предполагалось, что ставрикозавр был близок к тем древнейшим ящерам, от которых путём дивергенции произошли обе группы ящеротазовых динозавров. Однако в 1984 году в Аризоне в Пеинтед Дезерт, США, были найдены остатки еще более древнего динозавра и он оказался настолько похожим на ранних прозауропод, что стало ясно, что группа прозауропод эволюционно древнее, чем ставрикозавры. Так как ставрикозавр был весьма древним динозавром, и известные ученым части его скелета имеют очень примитивное строение, то можно сделать вывод, что и недостающие части его скелета были столь же примитивны. Таким образом, мы можем реконструировать облик ставрикозавра и предположить, что стопа и кисть этого ящера были пятипалыми, то есть всё ещё находились на ранней стадии эволюции (по другим данным верхние конечности ящера были украшены четырьмя пальцами).
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, герреразавриды, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| 02:51 pm [industrialterro] [Link] |
Saturnalia Сатурналия - самый ранний из известных травоядных динозавров. Первоначально его считали ранним прозауроподом. Теперь установлено, что он более примитивен, чем прозауроподы, поэтому классифицируется как ранний завроподоморф. По трём почти полностью сохранившимся скелетам мы можем воссоздать внешний облик этого изящного животного с длинной шеей и длинным хвостом. Голова маленькая, зубы грубо зазубреные, приспособленые к размалыванию растительной пищи. Тело обтекаемое, ноги тонкие. Кости таза примитивные (этот род находится на границе отряда динозавров), но голеностопные кости такие же, как у современных ему травоядных динозавров. По происхождению он был очень близок прозауроподам и зауроподам. Своё имя сатурналия получила в честь римского праздника зимнего солнцестояния, когда и был найден его скелет. Saturnalia was originally named on the basis of three partial skeletons. The holotype, MCP 3844-PV, a well-preserved semi-articulated postcranial skeleton, was discovered in mid-summer at Sanga da Alemoa, Rio Grande do Sul, in Brazil, in the geopark of Paleorrota. The two paratypes are MCP 3845-PV, partial skeleton including natural cast of partial mandible with teeth and some postcranial remains, and MCP 3846-PV, partial skeleton including postcranial remains. All specimen were collected in the "Wald-Sanga" (also known as "Sanga do Mato") locality from the Alemoa Member of the Santa Maria Formation (Rosário do Sul Group), dating to the Carnian faunal stage of the early Late Triassic, about 225 million years ago. A partial femur from the Carnian Pebbly Arkose Formation of Zimbabwe was also attributed to the genus. It is one of the oldest true dinosaurs yet found. It probably grew to about 1.5 meters (5 ft) long. The primitive nature of Saturnalia, combined with its mixture of sauropodomorph and theropod characteristics, has made it difficult to classify. Paleontologist Max Cardoso Langer and colleagues, in their 1999 description of the genus, assigned it to the Sauropodomorpha. However, in a 2003 paper, Langer noted that features of its skull and hand were more similar to the sister group of sauropodomorphs, the theropods, and that Saturnalia could at best be considered a member of the sauropodomorph "stem-lineage", rather than a true member of that group. José Bonaparte and colleagues, in a 2007 study, found Saturnalia to be very similar to the primitive saurischian Guaibasaurus. Bonaparte placed the two in the same family, Guaibasauridae. Like Langer, Bonaparte found that these forms may have been primitive sauropodomorphs, or an assemblage of forms close to the common ancestor of the sauropodomorphs and theropods. Overall, Bonaparte found that both Saturnalia and Guaibasaurus were more theropod-like than prosauropod-like. However, all more recent cladistic analyses found it to be a very basal sauropodomorph, possibly guaibasaurid, as the family was found to nest in a basal position within Sauropodomorpha.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, тероподы, ящеротазовые |
| 02:36 pm [industrialterro] [Link] |
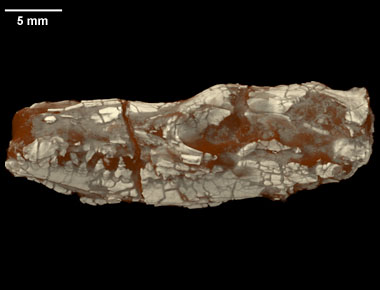
Sanjuansaurus Sanjuansaurus — род динозавров из семейства эрреразаврид, обнаруженный в 2010 году на территории современной Аргентины. Голотипом является экземпляр PVSJ 605. Представлен единственным видом — Sanjuansaurus gordilloi. Sanjuansaurus ("San Juan Province lizard") is a genus of herrerasaurid dinosaur from the Late Triassic-age Ischigualasto Formation of northwestern Argentina. It is known from and based on an associated and partially articulated partial skeleton (PVSJ 605) consisting of a jaw fragment, most of the vertebral column from the axis to the anterior tail, the shoulder blades, an ulna, part of the pelvis, most of the long bones of the legs, and a few other bones. PVSJ 605 was found in the Cancha de Bocas Member near the base of the Ischigualasto Formation, in Ischigualasto Provincial Park in San Juan. This horizon dates to approximately 231.4 Ma, during the late Carnian Stage of the Late Triassic. Sanjuansaurus was named and described in 2010 by Oscar Alcober and Ricardo Martinez. The type species is S. gordilloi after Raul Gordillo, the head fossil preparator and artist of the San Juan Museum. Sanjuansaurus was a contemporary of other early dinosaurs including Chromogisaurus, Eoraptor, Herrerasaurus, and Panphagia. It was comparable in size to a medium-sized Herrerasaurus, with a thigh bone length of 395 millimetres (15.6 in). Alcober and Martinez performed a phylogenetic analysis and found Sanjuansaurus to be a herrerasaurid.
Размеры тела в сравнении с человеком:
Обломок челюсти вверху, нижний цельный череп принадлежит герреразавру.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, герреразавриды, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| 02:18 pm [industrialterro] [Link] |
Riojasaurus Riojasaurus (meaning "Rioja lizard") was a herbivorous prosauropod dinosaur named after La Rioja Province in Argentina where it was found by José Bonaparte. It lived during the Late Triassic and grew to about 10 metres (33 ft) long. Riojasaurus is the only known riojasaurid to live in South America. Riojasaurus had a heavy body, bulky legs, and a long neck and tail. Its leg bones were dense and massive for a prosauropod. By contrast, its vertebrae were lightened by hollow cavities, and unlike most prosauropods, Riojasaurus had four sacral vertebrae instead of three. It probably moved slowly on all fours and was unable to rear up on its back legs. The nearly equal length of the fore and hindlimbs is also suggestive of an obligatorily quadrupedal gait. No skull was found with the first skeleton of Riojasaurus, although a well-preserved skull attributed to Riojasaurus was found later. The teeth of Riojasaurus were leaf shaped and serrated. The upper jaw contained 5 teeth at the front, with 24 more behind them in a row that ended under the eyes. Comparisons between the scleral rings of Riojasaurus and modern birds and reptiles suggest that it may have been cathemeral, active throughout the day at short intervals. Many scientists think that Riojasaurus was closely related to Melanorosaurus, the largest prosauropod known from the Triassic-Early Jurassic period. However, studies at Bristol University, England, suggest that it is unique in some key ways, such as the longer bones in its neck. It is certainly quite different from other prosauropods found in the Los Colorados Formation of Argentina. Due to their size and limb anatomy Riojasaurus and the possibly related Melanorosaurus have been considered close relatives of the earliest sauropods. However, if the hypothesis proposed by Peter Galton and Paul Sereno is correct -- namely, that prosauropods and true sauropods share a common ancestor rather than the former giving rise to the latter -- then commonalities shared by Riojasaurus and true sauropods are likely due to convergent evolution.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, прозавроподы, ящеротазовые |
| 01:53 pm [industrialterro] [Link] |
Procompsognathus Procompsognathus ( The name is derived from Compsognathus meaning 'elegant jaw' (Greek kompsos/κομψος meaning "elegant", "refined" or "dainty" and gnathos/γναθος meaning "jaw"), which was a later (Jurassic) dinosaur. The prefix προ/pro implies "before" or "ancestor of", although this direct lineage is not supported by subsequent research. Procompsognathus may have been about 1 meter long (3.3 ft). A biped, it had long hind legs, short arms, large clawed hands, a long slender snout with many small teeth, and a stiff tail. It lived in a relatively dry, inland environment and may have eaten insects, lizards, and other small prey. While it is undoubtedly a small, bipedal carnivore, the extremely poor preservation of the only known Procompsognathus fossil makes its exact identity difficult to determine. It has historically been considered a theropod dinosaur, though some, such as Allen (2004), have found Procompsognathus to be a primitive, non-dinosaurian ornithodiran. Sereno and Wild (1992) stated that the holotype specimen consisted of fossils from two separate animals. They referred the skull to the primitive crocodylomorph Saltoposuchus, and the remainder of the skeleton to a coelophysoid related to Segisaurus. Rauhut and Hungerbuhler (2000) noted features of the vertebrae which suggest that Procompsognathus may be a coelophysid or ceratosaur, and Carrano et al. (2005), in their re-study of the related genus Segisaurus, found both Segisaurus and Procompsognathus to belong to the Coelophysidae within Dinosauria.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, целофизиды, ящеротазовые |
| 01:19 pm [industrialterro] [Link] |
Pisanosaurus Pisanosaurus (meaning "Pisano lizard") is a genus of primitive ornithischian dinosaur from the Late Triassic of what is now South America. It was a bipedal herbivore described by Argentine paleontologist Rodolfo Casamiquela in 1967. Only one species, the type, Pisanosaurus mertii, is known, based on a single partial skeleton. The fossils were discovered in Argentina's Late Triassic Ischigualasto Formation, which dates to about 228 to 216.5 million years ago. The exact classification of Pisanosaurus has been the topic of debate by scientists for over 40 years; the current consensus is that Pisanosaurus is the oldest known ornithischian, part of a diverse group of dinosaurs which lived during nearly the entire span of the Mesozoic Era. Based on the known fossil elements, Pisanosaurus was a small, lightly built dinosaur approximately 1 m (3 ft 3 in) in length. Its weight was between 2,27–9,1 kg (5–20 lb). These estimates vary due to the incompleteness of the fossil. The tail of Pisanosaurus has been reconstructed as being as long as the rest of the body, based on other early ornithischians, but as a tail has not been recovered, this is speculative. It was bipedal and, like all other known ornithischians, was probably herbivorous. Pisanosaurus mertii was described by Argentine paleontologist Rodolfo Casamiquela in 1967. The name Pisanosaurus honors Juan A. Pisano, an Argentine paleontologist, while saurus is derived from the Greek σαυρος, meaning "lizard". Pisanosaurus is known from a single fragmented skeleton found in Argentina. It is based on a specimen given the designation PVL 2577, which was discovered in the Ischigualasto Formation. Pisanosaurus is very basal within Ornithischia; the postcrania seem to lack any good ornithischian synapomorphy; it was even suggested by Paul Sereno in 1991 that the fossil is a chimera. However, recent studies suggest that the fossils belong to a single specimen. Pisanosaurus has been classified either as a heterodontosaurid or as the earliest known ornithischian. A 2008 study placed Pisanosaurus outside of (and more basal than) Heterodontosauridae. In this study, Pisanosaurus is the earliest and most primitive ornithischian. Pisanosaurus is the type genus of the Pisanosauridae, a family erected by Casamiquela in the same paper which named Pisanosaurus. The Pisanosauridae family has fallen into disuse, as a 1976 study considered the group synonymous with the already named Heterodontosauridae.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, птицетазовые |
| March 23rd, 2012 | |
| 04:45 pm [industrialterro] [Link] |
Pantydraco Pantydraco — род ящеротазовых динозавров из группы Sauropodomorpha, живших в позднем триасовом периоде (около 228-199 миллионов лет назад), на территории нынешней Европы. Окаменелости были найдены в Англии. Впервые описан палеонтологом Yates и Kermack в 2007 году. Представлен одним видом - Pantydraco caducus. Pantydraco (where "panty-" is short for Pant-y-ffynnon, referring to the quarry in South Wales where it was found) was a genus of basal sauropodomorph dinosaur from the Late Triassic of the United Kingdom. It is based on a partial juvenile skeleton once thought to belong to Thecodontosaurus. In 2003, Adam Yates named the new species Thecodontosaurus caducus for a skull and partial skeleton (neck, partial shoulder girdle, and humeri) of a juvenile dinosaur, with additional material known for it. This material had been known in the scientific literature since 1984, and had been used to represent the genus Thecodontosaurus. However, changed understanding of the relationships and characteristics of basal sauropodomorphs (also known as prosauropods) has led Peter Galton, Yates, and D. Kermack to give T. caducus its own genus. As a basal sauropodomorph, Pantydraco would have been an omnivore, probably mostly bipedal.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, ящеротазовые |
| 04:03 pm [industrialterro] [Link] |
Panphagia Panphagia is a genus of sauropodomorph dinosaur described in 2009. It lived 231 million years ago, during the late Ladinian age of the mid Triassic period (according to the ICS; alternately the early Carnian age of the Late Triassic in the system used by the Geological Society of America), in what is now northwestern Argentina. The name Panphagia comes from the Greek words pan, meaning "all", and phagein, meaning "to eat", in reference to its inferred omnivorous diet. Panphagia is one of the earliest known dinosaurs, and is an important find which may mark the transition of diet in early sauropodomorph dinosaurs. Fossils of Panphagia were found in late 2006 by the Argentinean paleontologist Ricardo N. Martínez in rocks of the Ischigualasto Formation of Valle Pintado, Ischigualasto Provincial Park, San Juan Province, Argentina. The bones were found at approximately the same level as a 231,4 million year old ash layer, indicating it lived during the early Carnian of the Late Triassic. Panphagia is currently known from holotype PVSJ 874, the disarticulated remains of one partially grown individual of about 1.30 metres (4.3 ft) long. Portions of the skull, vertebrae, pectoral girdle, pelvic girdle, and hindlimb bones have been recovered. The russet-colored fossils were embedded in a greenish sandstone matrix and took several years to prepare and describe. Panphagia was described in 2009 by Ricardo N. Martínez and Oscar A. Alcober, both of the Museo de Ciencias Naturales, in San Juan, Argentina. They performed a phylogenetic analysis and found it to be the most basal known sauropodomorph dinosaur: the fossils shared similar features to those of Saturnalia, an early sauropodomorph, including similarities in the ischium, astralagus, and the scapular blade. Yet the fossils also exhibited similar features to those of Eoraptor, an early carnivorous saurischian, including hollow bones, sublanceolate teeth, and overall proportions. Based on analysis and comparison of the Panphagia fossils and those of its closest kin, Martínez and Alcober concluded that the evolution of saurischian dinosaurs likely began with small, cursorial animals similar to Panphagia, and that there is a "general similarity among all of these basal dinosaurs [suggesting] that few structural changes stand" between Panphagia, Eoraptor, and two basal theropods which have yet to be described. The type species of Panphagia is P. protos; the specific name, meaning "the first" in Greek, is a reference to its basal position. The teeth of Panphagia indicate a possible omnivorous diet, transitional in form between the mostly carnivorous theropods and the herbivorous sauropodomorphs. The teeth in the back of the jaw are shorter than those in the front, are leaf-shaped, and also have more marked serrations.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, ящеротазовые |

































































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}