[Recent Entries][Archive][Friends][User Info]
| April 21st, 2015 | |
|---|---|
| 03:50 pm [industrialterro] [Link] | Alectrosaurus (/əˌlɛktrɵˈsɔrəs/; meaning "alone lizard") is an extinct genus of tyrannosaurid theropod dinosaur that lived approximately 83 to 74 million years ago during the latter part of the Cretaceous Period in what is now Inner Mongolia. It was a medium sized, moderately-built, ground-dwelling, bipedal carnivore, with a body shape similar to its much larger relative, Tyrannosaurus rex, and could grow up to an estimated 5 m (16.4 ft) long. The generic name Alectrosaurus can be translated as "alone lizard," and is derived from the Greek words alektros and sauros ("lizard"). The meaning of the generic name is troublesome because the Greek work "alektros" (αλεκτρος) has also been alternatively translated as "unmarried" and even "rooster". At the time of its discovery, it was unlike any other Asian carnivore known, which may suggest that Gilmore selected a Greek word that reflected the meaning of alone. There is one named species (A. olseni), which is named in honor of George Olsen, who discovered the first specimens. Both genus and species were described and named by American paleontologist Charles Gilmore in 1933. In 1923, the Third Asiatic Expedition of the American Museum of Natural History, led by chief paleontologist Walter W. Granger was hunting for dinosaur fossils in Mongolia. On April 25th, assistant paleontologist George Olsen recovered the holotype (AMNH 6554), or name-bearing specimen, of Alectrosaurus, a nearly complete right hindlimb. This included the distal end of the right femur, the tibia, the fibula, the astragalus, the calcaneum, an incomplete right pes, three metatarsals of the left hind foot, two manual unguals, a manus, and the distal end of the pubis known as the pubic foot. On May 4th, Olsen discovered AMNH 6368 approximately 30 meters away from his first find. This specimen included a right humerus, two incomplete manual digits, four fragmentary caudal vertebrae, and other poorly preserved material. These discoveries were made at the Iren Dabasu Formation in what is now the Inner Mongolia Autonomous Region (Nei Mongol Zizhiqu) of the People's Republic of China. The age of this geologic formation is not clear, but is commonly cited as the Campanian stage of the Late Cretaceous Period, about 83 to 72 million years ago. More material, including comparable hind limb material as well as skull and shoulder elements, has been referred to Alectrosaurus. These fossils were found in the Bayan Shireh Formation of Outer Mongolia, a formation which is also of uncertain age. It may possibly extend into the early Campanian, but recent estimates suggest it was deposited from Cenomanian through Santonian times. Iren Dabasu and Bayan Shireh dinosaur faunas are similar, but van Itterbeecka et al. claimed that the Iren Dabasu is probably Campanian-Maastrichtian in age and possibly correlated with the Nemegt Formation, so it is not surprising that a species of Alectrosaurus would be found there. Furthermore, several more partial skeletons may have been found in both Inner and Outer Mongolia. Alectrosaurus was a medium sized, moderately built carnivorous dinosaur. The length of its tibia and femur are very close, in contrast to the majority of other tyrannosauroids, where the tibia is longer. The hind foot (and ankle) are also closer in size to the tibia than in most other tyrannosauroids, where the hind foot is usually longer. In 1933 Charles Gilmore examined the available material and concluded that AMNH 6554 and AMNH 6368 were syntypes belonging to the same genus. He based this on his observation that the manual unguals from both specimens were morphologically similar. Observing similarities with the hindlimbs of specimen AMNH 5664 Gorgosaurus sternbergi, he classified this new genus as a "Deinodont", a term that is now considered equivalent to tyrannosaurid. Due to its fragmentary nature, there is presently very little confidence in restoring its relationships with other tyrannosauroids and many recent cladistic analyses have omitted it altogether. One study recovered Alectrosaurus at no less than eight equally parsimonious positions in a tyrannosauroid cladogram. Alectrosaurus was originally characterized as a long-armed theropod, but Mader and Bradley (1989) observed that the forelimbs (AMNH 6368) did not belong to this individual and assigned them to the segnosauridae. The remaining material, AMNH 6554 represents the hind limb of a true tyrannosauroid, and were assigned as the lectotype for Alectrosaurus olseni. Mader and Bradley also described and assigned caudal vertebrae AMNH 21784 to this genus. These researchers concluded that Alectrosaurus was closely related to Maleevosaurus novojilov based on hind limb proportions. Some paleontologists have considered Alectrosaurus olseni to be a species of Albertosaurus. The Bayan Shireh material may or may not belong to this genus, and needs further study. One cladistic analysis showed that the two sets of specimens group together exclusive of any other taxa, so they are probably at least closely related, if not the same species. Alectrosaurus был двуногим плотоядным динозавром и по форме тела очень походил на своего гораздо большего собрата тираннозавра. Ящер был более миниатюрным, но очень быстрым, подвижным и успешным хищником. Длина его тела составляла от пяти до шести метров, высота 2-2,5 метра и вес от 450 килограммов до полутора тонн. У алектрозавра был довольно длинный череп, достигающий в длину примерно 60 сантиметров. Задний участок верхней челюсти длинной сто пятьдесят два миллиметра, высотой около пятидесяти миллиметров, толщиной двадцать три миллиметра имел шесть полых альвеол и одну неполную. Заднее окончание было обломано, однако место слома расположено близ анатомического окончания кости. Сохранившийся участок кости необычно низкий в сравнении с анатомическими участками у более поздних тираннозаврид. Судя по сохранившемуся фрагменту, число зубов у этого теропода из Джиракудука было большим, чем у тираннозавра и тарбозавра, но оно было сопоставимым с количеством зубов у примитивных мегалозаврид. Заднее предорбитальное отверствие простиралось вперед значительно дальше шестого зуба (при счёте от затылка). Примерно до этого уровня и практически строго по середине верхней кромки тянется глубокая борозда контакта с передним отростком скуловой кости. Четвертая альвеола сзади имеет размеры около 16 х 10 миллиметров. Изнутри невысоко над краем альвеол, тянется бороздка шириной полтора миллиметра, в которой располагаются отверстия для миграции зачатков сменных зубов внутрь челюсти. Рельеф этой области намного более гладкий, чем у тарбозавра, гребень на контакте с задней частью небной кости также с более гладким рельефом. Зубы, значительно уплощенные с боков, примерно как у дриптозавра, ножевидные, короткие и очень острые. Уплощенность с боков выражена намного сильнее, чем у тарбозавра и тем более у тираннозавра. Зубчики кромок мелкие с короткими слаборельефными насечками эмали, расположенными на их границах. По-видимому, латеральная уплощенность зубов, общая для позднемелового алектрозавра и позднеюрского торвозавра, свидетельствует не о близком родстве этих видов, а о сходном характере питания. Возможно, узкими ножеподобными зубами эти ящеры срезали мягкие ткани с костей, тогда как тираннозавриды кампана и маастрихта с помощью более толстых зубов могли отрывать крупные куски добычи даже с некрупными костями. Бедренные кости, предположительно относящиеся к алектрозавру, были с умеренно возвышающейся головкой. Нижнемедиальная часть головки имеет полушаровидную форму, в самом низу поверхность сустава отделена от несуставного участка кости невысокой ступенькой. Малый трохантер достаточно типичной для карнозавров формы – в виде субвертикального гребня, слабо выступающего вперед от поверхности кости. При взгляде на кость спереди он имеет вид небольшого, очень острого треугольника, ориентированного вершиной к верху. Четвертый трохантер, в виде узкого гребня, выступает сильнее, чем у тарбозавров. Передне-нижняя часть бедренной кости имеет лишь едва заметную экстенсорную бороздку. Когтевые фаланги передней и задней конечности относительно короткие и небольшие. Вообще одной из особенностей алектрозавра были тонкие ноги. Алектрозавр, является наиболее генерализованным тираннозавридом, но не предком более поздних представителей семейства. Если это так, а также если в Джиракудуке (местечко в Узбекистане, где найдено множество останков динозавров) действительнонайдены останки этого вида, то вытеснение (близ рубежа яруса коньяка и сантона) алектрозавров тарбозаврами в комплексах приморских низменностей, могла сложиться не за счет формирования последнего рода на прежней территории расселения алектрозавров, а за счет миграции конкурентоспособных тарбозавров из другого формообразовательного центра. Большое значение для установления филогенетической принадлежности среднеразмерных тиранозаврид позднего альба, раннего секомана и раннего турона имели бы новые сборы. Прошло более полувека с тех пор как начались планомерные исследования останков вымерших наземных позвоночных на территории Монгольской Народной Республики. Нахождение богатых захоронений в центре Азиатского материка открывало новые широкие перспективы. Первые находки монгольских динозавров уже носили многообещающий характер в плане их своеобразия и ожидаемых перспектив. Пожалуй, с самого начала палеонтологического исследования Центральной Азии и, в особенности, Монголии довольно ясно намечались контуры наиболее богато и разнообразно представленных групп динозавров, среди которых поздним группам хищных теропод ныне принадлежит бесспорное первенство. С единичных находок началось изучение монгольских хищных динозавров. Несовершенный по сохранности материал первых сборов уже тогда свидетельствовал, что исследователи стоят на пороге интересных открытий. Понадобились многие годы и усилия перечисленных крупных экспедиций, чтобы подготовить условия для дальнейшего продвижения вперед на действительно широкой и полноценной палеонтологической основе. Алектрозавр был обнаружен на территории автономного региона на севере КНР (Внутренняя Монголия) в 1923 году в отложениях кампанского яруса (формация Ирен-Дабасу). Название происходит от греческого слова «алектрос», что означает «одинокий» и «заурус» - «ящерица», что можно перевести как «Одинокий ящер». Первые образцы древнего ящера были обнаружены в 1923 году третьей по счету экспедицией снаряженной в Центральную Азию для очередных раскопок. Экспедицией состоящей из палеонтологов Нью-Йоркского Американского музея естественной истории во главе с Джорджем Олсеном и были обнаружены останки неизвестного на то время древнего ящера. В честь своего первооткрывателя Джорджа Олсена динозавр и получил свое имя – «Alectrosaurus olseni». Одновременно с находкой останков алектрозавра было обнаружено множество других костей динозавров. Ученые объясняют массовую находку тем, что в 1909 году здесь произошло сильнейшее наводнение, которое и раскрыло миру большое количество ископаемого материала. Были найдены частичный скелет и отдельные кости нескольких экземпляров алектрозавров. Кости алектрозавров несколько отличались от костей ранее известных азиатских хищных динозавров и являлись неизвестным и не изученным материалом. Обнаружив находку, ученые-палеонтологи, по мере ее изучения допустили ряд грубых ошибок и кости алектрозавров были перепутаны с костями других теропод, в частности, сегнозавров (семейство терезинозавров), найденных ранее на этой же территории. Обнаруженной находке учеными был присвоен номенклатурный номер или голотип ( AMNH 6554) состоящий из фрагментов верхней челюсти, зубов, бедренных костей, позвонков, тарсуса и когтевых фаланг. Путаница возникла из-за того, что ученые предположили, что алектрозавр обладал длинными и мощными передними конечностями, однако позднее выяснилось, что найденные рядом с остальными костями «руки» принадлежат совсем другому животному - неизвестному представителю терезинозавров, а передние конечности алектрозавра, как и у всех тираннозавров, были крошечными. При дальнейшем изучении и сопоставлении останков ученые-палеонтологи отнесли найденные задние конечности, фрагменты черепа и плечевые элементы к Alectrosaurus. Оставшийся материал, состоящий из задних конечностей истинного тираннозавроида, характеризуется низким соотношением между длиной голени и бедра, а это означает, что обе кости были примерно одинакового размера, в отличие от большинства других тираннозавроидов, где голень была, как правило, больше. Ступня (и лодыжки), также ближе по размерам к голени, чем у большинства тираннозавроидов. Из-за неполноты ископаемого материала, Alectrosaurus изучен плохо. По В рамках одной гипотезы Alectrosaurus является близким родственником альбертозавров, но не все ученые разделяют это мнение, а потому данное предположение остаётся недоказанным. Единственное, что можно сказать с уверенностью это то, что Alectrosaurus был хищником из надсемейства тираннозавроидов, куда входит и знаменитый тираннозавр . На сегодняшний день этот ящер известен лишь по отдельным частям скелета, полного скелета алектрозавра так и не было найдено. Палеонтологи всего мира надеются, что удача им все-таки улыбнется, и они смогут больше узнать и подробнее изучить алектрозавра за счет дальнейших открытий и находок. A diagnosis is a statement of the anatomical features of an organism (or group) that collectively distinguish it from all other organisms. Some, but not all, of the features in a diagnosis are also autapomorphies. An autapomorphy is a distinctive anatomical feature that is unique to a given organism. According to Carr (2005), Alectrosaurus can be distinguished based on the following characteristics:
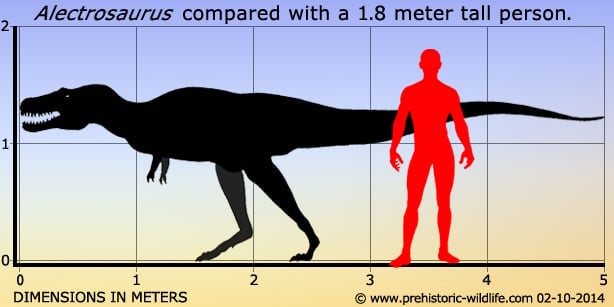
Alectrosaurus and Gigantoraptor were the top predators of their paleoenvironment and preyed on ornithischians like Bactrosaurus and Gilmoreosaurus. In a 2001 study conducted by Bruce Rothschild and other paleontologists, 23 foot bones referred to Alectrosaurus were examined for signs of stress fracture, but none were found. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
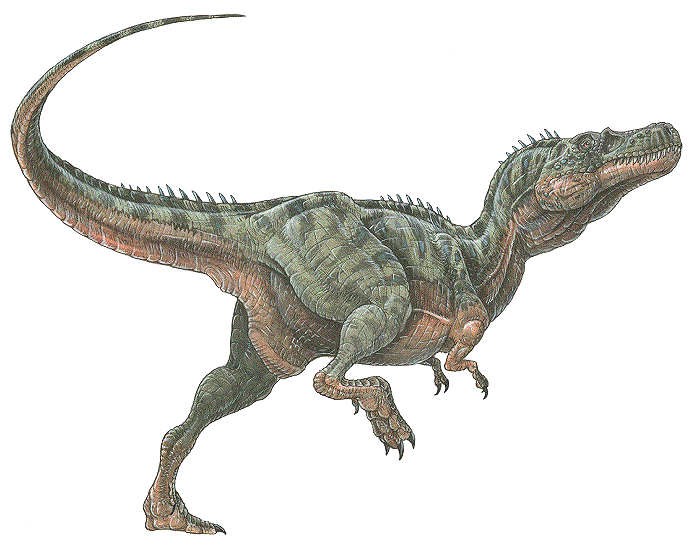





{kind=link}
{kind=link}