[Recent Entries][Archive][Friends][User Info]
Below are the 12 most recent journal entries recorded in the "Сообщество, посвящённое ра" journal:| April 24th, 2015 | |
|---|---|
| 01:26 pm [industrialterro] [Link] |
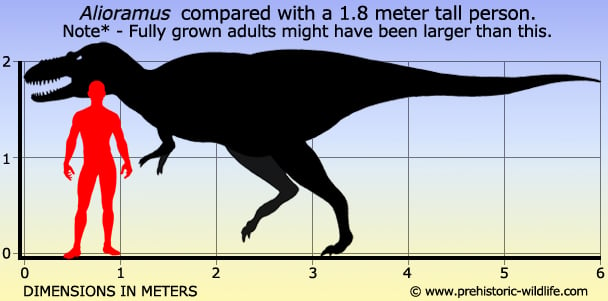
Alioramus Алиорам (Alioramus, от лат. alius ramus «другая ветвь») — род хищных динозавров из семейства тираннозавридов, живших в меловом периоде (около 70—65 млн лет назад) на территории современной пустыни Гоби (Монголия). Известны два вида. Впервые алиорама (A. remotus) описал советский палеонтолог С. М. Курзанов в 1976 году, обнаружив в монгольских отложениях фрагменты черепа и три плюсневые кости, принадлежащие динозавру, который жил 70-65 млн лет назад. Название от лат. alius ramus «другая ветвь» было выбрано из-за анатомических отличий алиорама от других тираннозаврид. С тех пор вопрос о принадлежности алиорама к тираннозавридам оставался дискуссионным. Новые останки обнаружены в монгольской пустыне Гоби в 2001 году сотрудниками Американского музея естественной истории под руководством Марка Норелла и Майкла Новачека. Они, вероятно, принадлежат девятилетнему подростку, который вырос всего на 85 %, жил 85—65 млн лет назад, достигал в длину 5 метров и весил около 369 кг. В 2009 году данные останки окончательно отнесли к новому виду — Alioramus altai. Главное отличие алтайского алиорама — необычные рога. Помимо обычных для тираннозавридов надбровных коротких рогов у него имеется по два пятисантиметровых рога, торчащих из каждой щеки. Использовать их для борьбы невозможно, поэтому предполагается, что рога служили украшением для привлечения самок. Алиорам по сравнению тарбозавром обладал маленьким черепом, миниатюрными зубками и слабыми челюстными мышцами и занимал отличную от последнего экологическую нишу. Alioramus (/ˌæli.ɵˈreɪməs/; meaning 'different branch') is a genus of tyrannosaurid theropod dinosaurs from the Late Cretaceous period of Asia. The type species, A. remotus, is known from a partial skull and three foot bones recovered from Mongolian sediments which were deposited in a humid floodplain about 70 million years ago. These remains were named and described by Soviet paleontologist Sergei Kurzanov in 1976. A second species, A. altai, known from a much more complete skeleton, was named and described by Stephen L. Brusatte and colleagues in 2009. Its relationships to other tyrannosaurid genera are unclear, with some evidence supporting a hypothesis that Alioramus is closely related to the contemporary species Tarbosaurus bataar. Alioramus were bipedal like all known theropods, and their sharp teeth indicate that they were carnivores. Known specimens were smaller than other tyrannosaurids like Tarbosaurus bataar and Tyrannosaurus rex, but their adult size is difficult to estimate since both species are known only from juvenile or sub-adult remains. The recent discovery of Qianzhousaurus indicates that it belongs to a distinct branch of tyrannosaur. The genus Alioramus is characterized by a row of five bony crests along the top of the snout, a greater number of teeth than any other genus of tyrannosaurid, and a lower skull than other tyrannosaurids. Alioramus remotus was estimated at 5 to 6 metres (16 to 20 ft) in length when originally described by Sergei Kurzanov in 1976. Kurzanov, however, did not correct for lengthening of the skull by deformation during fossilization, which may indicate a shorter overall body length for this individual. If this specimen is a juvenile, then adult Alioramus would have reached greater lengths, but no confirmed adult specimens are known. The skull of A. remotus was approximately 45 centimetres (18 in) long. In general, it is long and low, a shape typical of more basal tyrannosauroids and juveniles of larger tyrannosaurids. The premaxillary bones at the tip of the snout in Alioramus remotus have not been found, but are taller than wide in all tyrannosauroids for which they are known. The nasal bones are fused and ornamented with a row of five irregular bony crests that protrude upwards from the midline, where the nasal bones are sutured together. These crests all measure more than 1 centimetre (0.39 in) tall. At the back of the skull there is a protrusion, called the nuchal crest, arising from the fused parietal bones, a feature shared with all tyrannosaurids. In Alioramus, the nuchal crest is greatly thickened, similarly to Tarbosaurus and Tyrannosaurus. Like the rest of the skull, the lower jaw of Alioramus was long and slender, another possible juvenile characteristic. As in Tarbosaurus, a ridge on the outer surface of the angular bone of the lower jaw articulated with the rear of the dentary bone, locking the two bones together and removing much of the flexibility seen in other tyrannosaurids. Other tyrannosaurids had four premaxillary teeth, D-shaped in cross section, on each side. Including 16 or 17 in each maxilla, and 18 in each dentary, Alioramus had 76 or 78 teeth, more than any other tyrannosaurid. The braincase of A. altai was intermediate between the basal theropod and avialan conditions. The rest of the skeleton of Alioramus remotus is completely unknown except for three metatarsals (bones of the upper foot), but the discovery of A. altai, which is known from substantially more complete remains, has shed light on the anatomy of the genus. Paleontologists have long classified Alioramus within the superfamily Tyrannosauroidea, but because its remains were for many years poorly known, a more precise classification had remained elusive until the discovery of A. altai. A cladistic analysis published in 2003 found Alioramus could be further classified into the family Tyrannosauridae and the subfamily Tyrannosaurinae, alongside Tyrannosaurus, Tarbosaurus and Daspletosaurus. A 2004 study supported this result but suggested it was equally probable that Alioramus belonged outside the family Tyrannosauridae entirely, with its supposed juvenile characters actually reflecting a more basal position within Tyrannosauroidea. Another study omitted Alioramus altogether due to the only specimen's fragmentary nature. The discovery of A. altai in 2009 confirmed the placement of the genus within the Tyrannosaurinae. Tarbosaurus and Alioramus shared several skull features, including a locking mechanism in the lower jaw between the dentary and angular bones, and both lacked the prong of the nasal bones which connected to the lacrimal bones in all other tyrannosaurids except adult Daspletosaurus. The two genera may be closely related, representing an Asian branch of the Tyrannosauridae. Some specimens of Tarbosaurus have a row of bumps on the nasal bones like those of Alioramus, although much lower. The long and low shape of the only known Alioramus remotus skull indicated that it was immature when it died and might even have been a juvenile Tarbosaurus, which lived in the same time and place. The more prominent nasal crests and much higher tooth count of Alioramus, however, suggested it was a separate taxon, even if it is known only from juvenile remains, confirmed by the discovery of A. altai. Specimens identified as immature Tarbosaurus have the same tooth count as adults. The holotype (PIN 3141/1) of Alioramus is a partial skull associated with three metatarsals. A joint Soviet-Mongolian expedition to the Gobi Desert in the early 1970s found these remains at a locality known as Nogon-Tsav in the Mongolian province of Bayankhongor. Alioramus was named and described by Russian paleontologist Sergei Kurzanov in 1976. Its crests and low skull profile looked so different from other tyrannosaurids that Kurzanov believed his find was far removed from other members of the family. Accordingly, he gave it the generic name Alioramus, derived from the Latin alius ('other') and ramus ('branch'), and the specific name A. remotus, which means 'removed' in Latin. Alioramus is known from the holotypes of A. remotus and A. altai. The Beds of Nogon-Tsav are considered to be the same age as the Nemegt Formation. This geologic formation has never been dated radiometrically, but the fauna present in the fossil record indicate it was probably deposited during the Maastrichtian stage, at the end of the Late Cretaceous. The Maastrichtian stage in Mongolia, as preserved in the Nemegt Formation and at Nogon-Tsav, was characterized by a wetter and more humid climate compared with the semi-arid environment preserved in the earlier, underlying Barun Goyot and Djadochta Formations. Nemegt sediments preserve floodplains, large river channels and soil deposits, but caliche deposits indicate periodic droughts. This environment supported a more diverse and generally larger dinosaur fauna than in earlier times. Kurzanov reported that other theropods, including Tarbosaurus, ornithomimosaurs and therizinosaurs were discovered at the same locality, but these remains have never been reported in detail. If the Nogon Tsav fauna was similar to that of the Nemegt Formation, troodontid theropods, as well as pachycephalosaurs, ankylosaurids and hadrosaurs would also have been present. Titanosaurian sauropods were also potential prey for predators in the Nemegt. ( Read More ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9): ( Read More ) Размеры тела в сравнении с человеком:
Ископаемые останки и реплики (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые |
| April 21st, 2015 | |
| 03:50 pm [industrialterro] [Link] |
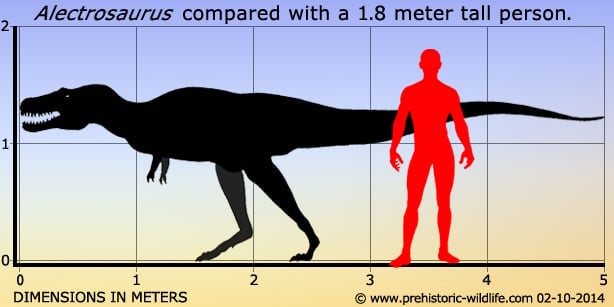
Alectrosaurus Alectrosaurus (/əˌlɛktrɵˈsɔrəs/; meaning "alone lizard") is an extinct genus of tyrannosaurid theropod dinosaur that lived approximately 83 to 74 million years ago during the latter part of the Cretaceous Period in what is now Inner Mongolia. It was a medium sized, moderately-built, ground-dwelling, bipedal carnivore, with a body shape similar to its much larger relative, Tyrannosaurus rex, and could grow up to an estimated 5 m (16.4 ft) long. The generic name Alectrosaurus can be translated as "alone lizard," and is derived from the Greek words alektros and sauros ("lizard"). The meaning of the generic name is troublesome because the Greek work "alektros" (αλεκτρος) has also been alternatively translated as "unmarried" and even "rooster". At the time of its discovery, it was unlike any other Asian carnivore known, which may suggest that Gilmore selected a Greek word that reflected the meaning of alone. There is one named species (A. olseni), which is named in honor of George Olsen, who discovered the first specimens. Both genus and species were described and named by American paleontologist Charles Gilmore in 1933. In 1923, the Third Asiatic Expedition of the American Museum of Natural History, led by chief paleontologist Walter W. Granger was hunting for dinosaur fossils in Mongolia. On April 25th, assistant paleontologist George Olsen recovered the holotype (AMNH 6554), or name-bearing specimen, of Alectrosaurus, a nearly complete right hindlimb. This included the distal end of the right femur, the tibia, the fibula, the astragalus, the calcaneum, an incomplete right pes, three metatarsals of the left hind foot, two manual unguals, a manus, and the distal end of the pubis known as the pubic foot. On May 4th, Olsen discovered AMNH 6368 approximately 30 meters away from his first find. This specimen included a right humerus, two incomplete manual digits, four fragmentary caudal vertebrae, and other poorly preserved material. These discoveries were made at the Iren Dabasu Formation in what is now the Inner Mongolia Autonomous Region (Nei Mongol Zizhiqu) of the People's Republic of China. The age of this geologic formation is not clear, but is commonly cited as the Campanian stage of the Late Cretaceous Period, about 83 to 72 million years ago. More material, including comparable hind limb material as well as skull and shoulder elements, has been referred to Alectrosaurus. These fossils were found in the Bayan Shireh Formation of Outer Mongolia, a formation which is also of uncertain age. It may possibly extend into the early Campanian, but recent estimates suggest it was deposited from Cenomanian through Santonian times. Iren Dabasu and Bayan Shireh dinosaur faunas are similar, but van Itterbeecka et al. claimed that the Iren Dabasu is probably Campanian-Maastrichtian in age and possibly correlated with the Nemegt Formation, so it is not surprising that a species of Alectrosaurus would be found there. Furthermore, several more partial skeletons may have been found in both Inner and Outer Mongolia. Alectrosaurus was a medium sized, moderately built carnivorous dinosaur. The length of its tibia and femur are very close, in contrast to the majority of other tyrannosauroids, where the tibia is longer. The hind foot (and ankle) are also closer in size to the tibia than in most other tyrannosauroids, where the hind foot is usually longer. In 1933 Charles Gilmore examined the available material and concluded that AMNH 6554 and AMNH 6368 were syntypes belonging to the same genus. He based this on his observation that the manual unguals from both specimens were morphologically similar. Observing similarities with the hindlimbs of specimen AMNH 5664 Gorgosaurus sternbergi, he classified this new genus as a "Deinodont", a term that is now considered equivalent to tyrannosaurid. Due to its fragmentary nature, there is presently very little confidence in restoring its relationships with other tyrannosauroids and many recent cladistic analyses have omitted it altogether. One study recovered Alectrosaurus at no less than eight equally parsimonious positions in a tyrannosauroid cladogram. Alectrosaurus was originally characterized as a long-armed theropod, but Mader and Bradley (1989) observed that the forelimbs (AMNH 6368) did not belong to this individual and assigned them to the segnosauridae. The remaining material, AMNH 6554 represents the hind limb of a true tyrannosauroid, and were assigned as the lectotype for Alectrosaurus olseni. Mader and Bradley also described and assigned caudal vertebrae AMNH 21784 to this genus. These researchers concluded that Alectrosaurus was closely related to Maleevosaurus novojilov based on hind limb proportions. Some paleontologists have considered Alectrosaurus olseni to be a species of Albertosaurus. The Bayan Shireh material may or may not belong to this genus, and needs further study. One cladistic analysis showed that the two sets of specimens group together exclusive of any other taxa, so they are probably at least closely related, if not the same species. ( Read More ) Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
| 01:02 pm [industrialterro] [Link] |
Albertosaurus (часть 1) Альбертоза́вр (Albertosaurus) — род теропод из семейства тираннозаврид. Существовал в западной части Северной Америки (от Аляски до Техаса) во время позднего мелового периода (76—65 миллионов лет назад). Ареал типового вида Albertosaurus sarcophagus был ограничен областью современной канадской провинции Альберты, отсюда и произошло его название. Как и все тираннозавриды, альбертозавр был двуногим хищником с крошечными двупалыми передними лапками и массивной головой с десятками больших острых зубов. Считается, что альбертозавр был доминирующим хищником в своей среде обитания. Будучи относительно крупным для теропода, по размерам он значительно уступал своему более известному родственнику тираннозавру — средний вес особей альбертозавра был менее двух тонн. Изучение вида ведётся с 1884 года, всего были обнаружены останки более тридцати особей, что позволило учёным изучить анатомию альбертозавра лучше других представителей семейства. Открытие двадцати особей различного возраста на одном участке (что может являться свидетельством стайного поведения) позволяет исследовать онтогенез и популяционную биологию альбертозавра, в отличие от большинства других динозавров. По размеру альбертозавр был меньше самых крупных тираннозаврид, например, тарбозавра и тираннозавра. Типичная взрослая особь достигала 9 метров в длину, некоторые старые особи могли превышать 10 метров. Вес взрослых альбертозавров, вычисленный на основе ископаемых останков разными методами, составлял от 1,2 до 1,7 тонны. Массивный череп альбертозавра, достигавший 1 метра в длину, поддерживала короткая S-образная шея. Широкие отверстия в черепе (окна) снижали вес головы, а также предоставляли пространство для челюстных мышц и сенсорных органов. Длинные челюсти альбертозавра содержали более 60 длинных, похожих на бананы зубов (более крупные тираннозавриды имели меньшее количество зубов). В отличие от большинства других теропод, альбертозавр и другие тираннозавриды были гетеродонтами, то есть имели разные виды зубов, в зависимости от их расположения во рту. Зубы, находившиеся на резцовой кости, были меньше остальных, располагались более тесно и имели D-образную форму в сечении. Над глазами имелись короткие костяные гребни, которые при жизни могли быть яркого цвета и использоваться при ухаживании для привлечения партнёра. Все тираннозавриды, в том числе альбертозавр, имеют сходное строение тела. Как и большинство тероподов, альбертозавр был бипедальным и уравновешивал тяжёлую голову и корпус очень длинным хвостом. Передние конечности верхнемеловых тираннозаврид были чрезвычайно мелкими и сохранили только два когтя. Задние конечности были длинными и заканчивались четырьмя пальцами с мощными когтями. Короткий первый палец задней конечности не участвовал в передвижении, только три других соприкасались с землёй, третий (средний) был длиннее остальных. Считается, что альбертозавр мог развивать скорость от 14 до 21 км/ч. Альбертозавр был представителем семейства тираннозаврид, подсемейства альбертозаврин. Его самый близкий «родственник» — обнаруженный в породах более позднего возраста горгозавр (иногда называемый Albertosaurus libratus). Эти два рода — единственные известные представители подсемейства альбертозаврин, но могли существовать и другие. Палеонтолог Томас Хольц в 2004 году предположил, что аппалачиозавр тоже относился к альбертозавринам, но в своей более поздней работе уже не рассматривал аппалачиозавра как представителя этого подсемейства, и эта точка зрения поддерживается и другими учёными. Другим подсемейством тираннозаврид являются тираннозаврины, к которым относятся дасплетозавр, тарбозавр и тираннозавр. По сравнению с тираннозавринами, альбертозаврины имели более стройное тело, пропорционально меньшие черепа и кости таза и более длинные большеберцовые и таранные кости, а также кости плюсны и пальцев. Название этому виду динозавров дал известный американский геолог, евгеник и палеонтолог Генри Осборн в 1905 году в одностраничном комментарии в конце своего описания Tyrannosaurus rex. Название «альбертозавр» происходит от названия канадской провинции Альберта, на территории которой впервые были найдены кости. Родовое название также включает в себя греческое слово σαῦρος (saurus) — ящер. Название типового вида «sarcophagus» означает «хищник», «плотоядный» и имеет то же происхождение, что и слово «саркофаг» (от древнегреческого σάρξ (sarx) — плоть и φαγεῖν (phagein) — поедать). Науке известно более тридцати скелетов этого динозавра всех возрастов. Типовым экземпляром является фрагментированный череп, обнаруженный в 1884 году в отложениях карьера Хорсшу одноимённого каньона, расположенного рядом с рекой Ред-Дир на территории современной провинции Альберта. Этот экземпляр и меньший череп, вместе с некоторым скелетным материалом, были обнаружены геологической экспедицией во главе с известным геологом Джозефом Тирреллом. Эти два черепа были отнесены к ранее описанному виду Laelaps incrassatus Эдвардом Коупом в 1892 году, несмотря на то, что название Laelaps было ранее дано роду клещей и было изменено на Dryptosaurus в 1877 году Чарлзом Маршем. Коуп отказался признать новое название, созданное его главным научным противником; несмотря на это, Лоуренс Ламбе в 1904 году описал эти черепа, отнеся их к виду Dryptosaurus incrassatus, а не Laelaps incrassatus. Чуть позже Генри Осборн указал на то, что описание Dryptosaurus incrassatus было основано на нескольких основных зубах тираннозаврид, поэтому два черепа из Альберты не могут с уверенностью быть отнесены к этому виду. Эти черепа также значительно отличались от останков типового вида дриптозавра — Dryptosaurus aquilunguis. Таким образом, Осборн в 1905 году выделил на основании этих черепов новый вид — Albertosaurus sarcophagus. При классификации он полагался на описание, созданное Ламбе на год раньше. Оба экземпляра (КМП 5600 и КМП 5601) были выставлены в Канадском музее природы в Оттаве. В 1910 году американский палеонтолог Барнум Браун обнаружил множество костей, принадлежавших большой группе альбертозавров в другом карьере, рядом с рекой Ред-Дир. Из-за большого количества образцов и недостатка времени экспедиция Брауна не откопала все экземпляры, но всё же ей удалось собрать из костеносного слоя все особи, которые можно было идентифицировать на тот момент. В тот год было обнаружено семь разных наборов костей стопы, а также два образца, несоответствующих другим по размерам и, возможно, принадлежащих детёнышам. Все они находятся в коллекции Американского музея естественной истории. Королевский Тиррелловский палеонтологический музей возобновил экспедицию в этот район (Сухой остров Буффало-Джамп) в 1997 году. Раскопки проводились с 1997 по 2005 год. В результате были обнаружены кости ещё 13 особей, среди них двухлетнего детёныша и очень старого альбертозавра, длина которого была оценена в 10 метров. Ни один скелет из найденных экземпляров не был полным. Кости были выставлены в Тирелловском музее и Американском музее естественной истории. В 1913 году палеонтолог Чарльз Штернберг обнаружил скелет тираннозаврида в формации Парк динозавров в Альберте. В следующем 1914 году Лоуренс Ламбе дал этому динозавру название Gorgosaurus libratus. Позже в Альберте и американском штате Монтана были найдены другие останки горгозавров. Обнаружив, что горгозавр и альбертозавр имеют слишком мало отличий, Дейл Рассел предложил считать горгозавра младшим синонимом альбертозавра, а так как альбертозавр был открыт и назван первым, в 1970 году горгозавр был переименован в Albertosaurus libratus, из-за чего временной диапазон обитания альбертозавра увеличился на несколько миллионов лет, а ареал простирался на сотни километров южнее, чем это считалось раньше. В 2003 году палеонтолог Фил Карри изучил черепа нескольких тираннозаврид, в том числе и горгозавра и альбертозавра, и выяснил, что они имеют всё же несколько больше различий, чем это считалось ранее. На сегодняшний день вопрос о том, использовать один или два рода, не решён окончательно, так как горгозавр и альбертозавр являются очень близкими таксонами. Признавая это, Карри, однако, рекомендовал, чтобы альбертозавр и горгозавр считались отдельными родами, поскольку отличий между ними не меньше, чем между дасплетозавром и тираннозавром, которых почти всегда можно отличить друг от друга. Кроме того, недавно были обнаружены новые скелеты альбертозаврин в штате Нью-Мексико и на Аляске, и Карри предположил, что вопрос о альбертозавре-горгозавре может быть полностью разъяснён, как только они будут изучены. Большинство исследователей поддерживают точку зрения Карри, но есть и противники этой гипотезы. В 1928 году Уильям Паркс описал новый вид — Albertosaurus arctunguis на основании частичного скелета, раскопанного у реки Ред-Дир, но с 1970 года этот вид считается идентичным Albertosaurus sarcophagus. Этот экземпляр (КМО 807) хранится в Королевском музее Онтарио в Торонто. С тех пор в Альберте было обнаружено ещё шесть экземпляров, причисленных к этому виду. Ископаемые, причисленные к Albertosaurus sarcophagus, были также обнаружены в американских штатах Монтана, Нью-Мексико и Вайоминг, но они, вероятно, не относятся к этому виду, а возможно, и к роду альбертозавр. Albertosaurus megagracilis был описан на основании небольшого скелета, найденного в геологической формации Хелл-Крик. В 1995 году он был выделен в отдельный род — Dinotyrannus, который позже стали описывать как принадлежащий подростковой особи тираннозавра рекса. Репродукции (1, 2, 3, 4, 5, 6):
Размеры тела в сравнении с человеком: Ископаемые останки и реплики (1, 2, 3, 4, 5, 6):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые |
| 12:50 pm [industrialterro] [Link] |
Albertosaurus (часть 2) До нашего времени сохранились останки почти всех возрастных групп альбертозавра. С помощью гистологического исследования можно определить возраст животного в момент его смерти. Сравнив останки особей одного вида, но разного возраста, можно определить среднюю скорость роста этих животных. Самый молодой из известных альбертозавров — двухлетний, он был обнаружен в Буффало-Джамп, весил приблизительно 50 килограммов, а в длину достигал чуть более двух метров. Десятиметровый экземпляр, обнаруженный в том же карьере, является самым старым из известных — он умер в возрасте двадцати восьми лет. Наибольший темп роста альбертозавра приходился на период от четырёх до шестнадцати лет, как и у некоторых других тираннозаврид. В этот период жизни альбертозавры набирали 122 килограмма ежегодно, пока не достигали 1,3 тонны. Впрочем, согласно другим исследованиям, вес взрослых альбертозавров был больше, и темпы роста тоже могли быть больше. У тираннозаврид, близких по размерам к альбертозавру, темпы роста были схожи, а более крупные, такие как тираннозавр, наоборот, росли почти в пять раз быстрее (601 килограмм ежегодно). Конец фазы быстрого роста у альбертозавров предполагал достижение половой зрелости; хотя они и продолжали расти до конца жизни, но гораздо медленнее. Раннее половое созревание в процессе роста типично для мелких и крупных динозавров, как и для большинства млекопитающих, таких как люди и слоны. С другой стороны, ближайшие родственники динозавров, птицы, достигают половой зрелости лишь по завершении процесса роста. Чаще всего встречаются останки особей альбертозавров в возрасте четырнадцати лет и старше. Более молодых животных находят редко по нескольким причинам — в основном из-за того что кости молодых животных сохраняются хуже, чем кости взрослых, а также из-за того что искателям окаменелостей их сложнее заметить. Несмотря на то что молодые альбертозавры были достаточно крупными животными, их останков обнаружено гораздо меньше, чем взрослых. Предполагается, что такая ситуация сложилась вследствие хороших условий жизни для детёнышей, из-за чего они умирали реже взрослых. Согласно общей теории, уже по достижении всего двух лет детёныши альбертозавра были крупнейшими хищниками своего ареала, они также были гораздо быстрее, чем все их жертвы. Из-за этого даже у молодых альбертозавров не было врагов. Это и привело к уменьшению смертности детёнышей и, соответственно, к уменьшению количества их останков. Останки двенадцатилетних альбертозавров встречаются в два раза чаще, чем более молодых, а достигших половой зрелости четырнадцатилетних и старше — ещё в два раза больше. Возможно, по достижении половой зрелости усиливалась внутривидовая борьба за самку или территорию. Более высокая смертность взрослых объясняет большее количество сохранившихся останков. Очень крупные особи также встречаются очень редко, поскольку лишь немногим удавалось прожить столь долго, чтобы достигнуть таких размеров. Схема, в которой высокий уровень смертности наблюдается среди детёнышей, низкий среди подростков, повышенный среди взрослых особей и крайне небольшое количество животных способно дожить до глубокой старости, свойственна большинству современных крупных животных, таких как носороги, африканский буйвол или слон, а также некоторым тираннозавридам, за исключением, разумеется, альбертозавра, детёныши которого умирали редко. В связи с пробелами в палеонтологической летописи, выводы об уровне смертности не могут быть более точными, в том числе и потому что более двух третей всех останков альбертозавров были найдены в одном месте. В Буффало-Джамп были обнаружены останки двадцати двух альбертозавров, что является самым крупным массовым захоронением тероподов после массового захоронения аллозавров в карьере динозавров Кливленд-Ллойд в Юте. В захоронении присутствуют останки одного старого взрослого, восьми взрослых между семнадцатью и двадцатью тремя годами, семи подростков в возрасте от двенадцати до шестнадцати лет и шести детёнышей между двумя и одиннадцатью годами, которые ещё не достигли фазы быстрого роста. Почти полное отсутствие останков травоядных животных и одинаковый уровень сохранности останков альбертозавров в карьере навели Фила Карри на мысль, что это захоронение не было ловушкой для хищников, как смоляные ямы на ранчо Ла-Брея в Калифорнии, и что все сохранившиеся животные умерли в одно время. Карри считает, что это свидетельство стайного поведения. Другие учёные более скептичны, отмечая, что, возможно, животных согнала вместе засуха или их тела были снесены в одно место наводнением. Есть многочисленные свидетельства стадного образа жизни среди растительноядных динозавров, в том числе цератопсов и гадрозавров. Однако массовые захоронения хищных динозавров редки. Маленькие тероподы, такие как дейноних, целофиз и мегапнозавр (синтарс), были найдены в больших скоплениях, как и крупные хищники аллозавр и мапузавр. Есть также доказательства стайного поведения среди других тираннозаврид. Фрагментарные остатки детёнышей были найдены рядом с тираннозавром по прозвищу «Сью», хранящимся в музее естественной истории имени Филда в Чикаго, а в костеносном слое геологической формации Ту-Медисин в Канаде были найдены останки трёх дасплетозавров рядом с несколькими гадрозаврами. Эти находки могут подтвердить, что альбертозавр также вёл социальный образ жизни, хотя некоторые вышеупомянутые скопления могут быть временными или неестественными. Но этому может быть и другое объяснение. По крайней мере, некоторые из этих находок представляют собой проявления массового каннибализма, как у комодского варана. Карри также считает, что доказательством стайного образа жизни альбертозавров может быть их способ охоты. Пропорции ног молодых альбертозавров были сопоставимы с таковыми у орнитомимид, которые, пожалуй, были самыми быстрыми среди динозавров. Они, по всей видимости, были быстрее любых травоядных своего времени. Карри выдвинул гипотезу, что младшие члены стаи отбивали жертву от стада и загоняли её к взрослым особям, которые были сильнее детёнышей, но медлительней. Животные подросткового возраста, предположительно, вели образ жизни, отличавшийся от поведения своих взрослых родственников, заполняя экологическую нишу между ними и мелкими тероподами, крупнейшие из которых весили вдвое меньше взрослого альбертозавра. Подобная ситуация наблюдается у современных комодских варанов, которые начинают жизнь как маленькие насекомоядные животные, а потом становятся доминирующими хищниками. Тем не менее, полностью воссоздать поведение вымерших животных на основании окаменелостей невозможно. Все идентифицируемые окаменелости Albertosaurus sarcophagus были обнаружены в формации каньона Хорсшу. Эта геологическая формация относится к маастрихтскому ярусу мелового периода, возрастом 70—73 миллиона лет. Непосредственно ниже этой формации находятся сланцы морской формации Бирпо, которая является частью более обширной морской формации Сиуэй. В конце мелового периода море отступало, климат становился холоднее, уровень моря уменьшался, обнажая землю, которая раньше находилась под водой. Этот процесс, однако, не был гладким: из-за сильных бурь на море часть суши в районе каньона Хорсшу частично затоплялась, но в конце концов море всё-таки отступило. Из-за постоянно изменяющегося уровня моря в формации Хорсшу было много прибрежных лагун, болот и илистых осушек. Многочисленные угольные пласты представляют древние торфяные болота. Как и в случае большинства других позвоночных в этой формации, останки альбертозавра были найдены в отложениях доисторических дельт и пойм больших рек. Доисторическая фауна формации Хорсшу хорошо известна, поскольку окаменелости позвоночных, включая динозавров, присутствуют здесь в массовых количествах. Фауну рыб составляли акулы, скаты, осетры, ильные рыбы, панцирникообразные и подобные панцирниковым аспидохиноморфы. Фауна млекопитающих включала в себя многобугорчатых и сумчатых, таких как дидельфодон. В морских отложениях был обнаружен плезиозавр леуроспондил, а в отложениях пресноводной среды были найдены черепахи, хампсозавры и крокодилы, такие как лейдиозух и стангерохампс. Доминирующими представителями фауны были динозавры, особенно распространены были гадрозавры, которые составляют половину всех найденных здесь динозавров, к ним относились эдмонтозавр, зауролоф и гипакрозавр. Цератопсы и орнитомимиды также были очень распространены, составляя треть найденных здесь динозавров. Наряду с намного более редко встречающимися анкилозаврами и пахицефалозаврами, эти растительноядные динозавры были добычей для множества плотоядных тероподов, таких как троодонтиды, дромеозавриды и кенагнатиды. Взрослые альбертозавры были доминирующими хищниками в этой среде и находились на вершине пищевой пирамиды, а промежуточные ниши между мелкими тероподами и взрослыми альбертозаврами, возможно, занимали юные альбертозавры. William Abler observed in 2001 that Albertosaurus tooth serrations resemble a crack in the tooth ending in a round void called an ampulla. Tyrannosaurid teeth were used as holdfasts for pulling meat off a body, so when a tyrannosaur pulled back on a piece of meat, the tension could cause a purely crack-like serration to spread through the tooth. However, the presence of the ampulla distributed these forces over a larger surface area, and lessened the risk of damage to the tooth under strain. The presence of incisions ending in voids has parallels in human engineering. Guitar makers use incisions ending in voids to, as Abler describes, "impart alternating regions of flexibility and rigidity" to the wood they work with. The use of a drill to create an "ampulla" of sorts and prevent the propagation of cracks through material is also used to protect aircraft surfaces Abler demonstrated that a plexiglass bar with incisions called "kerfs" and drilled holes was more than 25% stronger than one with only regularly placed incisions. Unlike tyrannosaurs, ancient predators like phytosaurs and Dimetrodon had no adaptations to prevent the crack-like serrations of their teeth from spreading when subjected to the forces of feeding. In 2009, researchers hypothesized that smooth-edged holes found in the fossil jaws of tyrannosaurid dinosaurs such as Albertosaurus were caused by a parasite similar to Trichomonas gallinae, which infects birds. They suggested that tyrannosaurids transmitted the infection by biting each other, and that the infection impaired their ability to eat food. In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. They found that only one of the 319 Albertosaurus foot bones checked for stress fractures actually had them and none of the four hand bones did. The scientists found that stress fractures were "significantly" less common in Albertosaurus than in the carnosaur Allosaurus. ROM 807, the holotype of A. arctunguis (now referred to A. sarcophagus), had a 2.5 by 3.5 cm deep hole in the iliac blade, although the describer of the species did not recognize this as pathological. The specimen also contains some exostosis on the fourth left metatarsal. In 1970, two of the five Albertosaurus sarcophagus specimens with humeri were reported by Dale Russel as having pathological damage to them. In 2010, the health of the Dry Island Albertosaurus assembly was reported upon. Most specimens showed no sign of disease. On three phalanges of the foot strange bony spurs, consisting of abnormal ossifications of the tendons, so-called enthesophytes, were present, their cause unknown. Two ribs and a belly-rib showed signs of breaking and healing. One adult specimen had a left lower jaw showing a puncture wound and both healed and unhealed bite marks. The low number of abnormalities compares favourably with the health condition of a Majungasaurus population of which it in 2007 was established that 19% of individuals showed bone pathologies. Тираннозаври́ды (Tyrannosauridae) — семейство тероподовых динозавров, вымерших в конце мелового периода (65 миллионов лет назад). Обитали в Азии и в Северной Америке. Известны с раннего мела. Подразделяются на два подсемейства — длинномордых Albertosaurinae и короткомордых Tyrannosaurinae. К тираннозавридам относят такие вымершие роды, как тарбозавр (длина 10 метров), дасплетозавр (длина 8,5 метров), монгольский алиорам (длина 6 метров), алтайский алиорам и другие. Самым известным представителем надсемейства является тираннозавр (занимает 4 место по размеру среди тероподов). Тираннозавр имел длину до 14 метров, в высоту достигал 5 метров, а весил около 7 тонн. Тираннозавриды главным образом характеризуются общими скелетными структурами, особенно характерны структуры бедренных и соединение носовых костей. Массивные головы и миниатюрные передние лапы с двумя пальцами делают этих меловых двуногих хищников легко узнаваемыми. Ранние тираннозавриды не имели характерного широкого, массивного черепа и редуцированных передних конечностей, как их потомки. Эти ранние виды обычно имели маленькие или средние черепа относительно размера их тела, были легче, стройнее, имели длинные, трёхпалые передние конечности. У них не было загнутых (бананоподобных) зубов, типичных для тираннозавровых. Внешне они наверняка были больше похожи на целурозавров чем на поздних тираннозавров. Некоторые исследователи предлагают, что некоторые или все известные «целурозавры» могут быть ранними тираннозавридами, хотя детальное изучение ещё не опровергло и не подтвердило эту теорию. Тираннозавриды — главным образом обитатели Евразии, а в Северную Америку они проникли лишь в конце мела, когда образовался сухопутный мост через Берингов пролив — Берингия. Об этом свидетельствуют находки костей в префектуре Исикава, Япония, которые предположительно принадлежат данному семейству и датированы возрастом около 140 млн лет. В Северной Америке тираннозавриды были найдены на территории Канады и США. В 2011 году был описан новый род Teratophoneus, найденный в штате Юта в слоях датируемых поздним мелом — 75 млн лет. Репродукции (7, 8, 9, 10, 11, 12): Размеры тела в сравнении с человеком: Ископаемые останки и реплики (7, 8, 9, 10, 11, 12): Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые |
| January 25th, 2013 | |
| 02:38 pm [industrialterro] [Link] |
Juratyrant "Juratyrant" (meaning "Jurassic tyrant") is a genus of tyrannosauroid dinosaur from the late Jurassic period (early Tithonian age) of England. The genus contains a single species, "J." langhami. The species is known from a single specimen consisting of an "associated partial skeleton represented by a complete pelvis" as well as a partially complete leg, and neck, back, and tail vertebrae. This skeleton was discovered in 1984 in Dorset. The specimen was mentioned in several papers, but was not formally described until 2008. The species was named in honor of commercial fossil collector Peter Langham, who uncovered the specimen. The specimen was discovered in strata of the Kimmeridge Clay dating from the Tithonian, the final stage of the Late Jurassic, and belonging to the Pectinatites pectinatus ammonite zone, indicating the fossil is between 149.3 and 149 million years old. The species was originally assigned to the genus Stokesosaurus, as Stokesosaurus langhami, by Roger Benson in 2008. However, later studies showed that it was not necessarily a close relative of Stokesosaurus clevelandi, the type species of that genus. It is set to be re-classified in its own genus, Juratyrant, in 2012 by Benson and Stephen Brusatte.
Размеры тела в сравнении с человеком:
Обозначен как (A). Обозначен как (А) и (В).
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
| January 21st, 2013 | |
| 05:40 pm [industrialterro] [Link] |
Tanycolagreus Tanycolagreus is a genus of coelurid theropod from the Late Jurassic of North America. In 1995 Western Paleontological Laboratories, Inc. uncovered the partial skeleton of a small theropod at the Bone Cabin Quarry West locality, Albany County, Wyoming, from the Salt Wash Member of the Morrison Formation, dating to the Oxfordian-Tithonian. At first the find was considered to be a specimen of Coelurus but subsequent study indicated it represented a species new to science, that in 2001 was announced to be named Tanycolagreus topwilsoni. It was actually named and described by Kenneth Carpenter, Clifford Miles and Karen Cloward in 2005. The etymology of the generic name Tanycolagreus, suggested by Ben Creisler, is based upon the greater length of its forelimbs and hindlimbs compared to Coelurus. It is derived from the Greek prefix τανυ~, tany~: 'long, stretched out', κῶλον, kolon: 'limb' and ἀγρεύς, agreus: 'hunter'. The specific name honours George Eugene "Top" Wilson, the father of a benefactor financially supporting the scientific research. The fossil, holotype TPII 2000-09-29, was donated to science by an anonymous benefactor. It is part of the collection of Thanksgiving Point Institute, Inc. and displayed in the North American Museum of Ancient Life at Lehi, Utah. It includes an incomplete skull and mandible (lower jaws) and much of the postcranial skeleton, i.e. the parts behind the head. The skull of Tanycolagreus is less well known than its postcranial anatomy, and only the following elements have been found: left nasal, left lacrimal, left premaxilla and one premaxillary tooth, left postorbital, left quadratojugal, incomplete left squamosal, right quadrate, right splenial, left articular, and two cheek teeth. A paratype has been assigned to the species: specimen AMNH 587 consisting of an incomplete hand also collected from Bone Cabin Quarry and originally in 1903 by Henry Fairfield Osborn referred to Ornitholestes hermanni. Two other fossils have been referred to Tanycolagreus: UUVP 2999, a premaxilla, originally in 1974 referred to Stokesosaurus clevelandi, from the Cleveland-Lloyd Quarry of Utah; and USNM 5737, a pair of distal pubes from Colorado earlier in 1920 by Charles Whitney Gilmore referred to Coelurus. These specimens are from the later Brushy Basin Member. Tanycolagreus is present in stratigraphic zone 2 of the Morrison. Remains possibly referrable to Stokesosaurus have been recovered from stratigraphic zone 5 of the Morrison Formation. A life restoration of Tanycolagreus is also on display at the North American Museum of Ancient Life, where it is portrayed as preying upon a small ornithischian dinosaur, Othnielia rex. Carpenter et al. (2005, pp. 43–44) determined that the holotype of Tanycolagreus represents a subadult individual which measured approximately 3.3 meters (11 ft) long in life. However, one of the referred fossils, the premaxilla from the Cleveland-Lloyd Quarry, would have belonged to a larger individual, measuring 4 meters (13 ft) long. In 2010 Gregory S. Paul estimated the weight of a four metres long animal at hundred twenty kilogrammes. It cannot be determined whether or not the Cleveland-Lloyd specimen represents a fully mature adult, so the upper size limit for the taxon remains unknown. The head of Tanycolagreus is large, elongated and rectangular in profile due to a blunt snout. The leg is rather long and lightly built. Carpenter et al. (2005; pp. 27 & 29) diagnosed Tanycolagreus topwilsoni as follows: "Medium-sized tetanuran having short, deep-bodied premaxilla pierced by narial foramen at base of nasal process, orbital process on postorbital, T-shaped quadratojugal, centrodiapophyseal lamina on dorsals. Differs from Coelurus in the absence of pleurocoel on anterior dorsals; posterior caudal prezygapophyses elongated to one-third centrum length, rather than short; straight, rather than sigmoidal, humeral shaft; bowed, rather than straight, radius; flat-bottomed rather than arced pubic foot; straight rather than sigmoidal femoral shaft; metatarsal length subequal to humeral length, rather than 1.75 times humeral length. Differs from Ornitholestes in straight anterior margin of premaxilla, rather than rounded; T-shaped rather than L-shaped quadratojugal; elongate neural spine; posterior caudal prezygapophyses only one-third centrum length, rather than one-half centrum length; bowed, slender radius, rather than straight, robust radius". The single premaxillary tooth preserved with the holotype is badly damaged, but does exhibit the asymmetrical cross-section typical in theropod teeth; the cheek teeth are too poorly preserved to show any detail. In the foot the second toe is slightly hyperextendable but does not carry an enlarged claw. Carpenter e.a. originally assigned Tanycolagreus to the Coeluridae. Carpenter et alii (2005, p. 44) state that, of the other known Morrison theropods, this genus most closely resembles Coelurus, though it retains more "primitive" features. A detailed phylogenetic analysis in 2007 by Philip Senter including Tanycolagreus showed it had a basal position in the Tyrannosauroidea. Later analyses indicated a basal position in the Coelurosauria.
Ископаемые останки и реплики (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целуриды, целурозавры, ящеротазовые |
| January 20th, 2013 | |
| 02:54 pm [industrialterro] [Link] |
Stokesosaurus Stokesosaurus (meaning "Stokes' lizard") is a genus of small (around three to four meters (10-13 ft) in length), carnivorous early tyrannosauroid theropod dinosaurs from the late Jurassic period of Utah. From 1960 onwards Utah geologist William Lee Stokes and his assistant James Madsen excavated thousands of disarticulated Allosaurus bones at the Cleveland-Lloyd Dinosaur Quarry in Emery County, Utah. During the early seventies Madsen began to catalogue these finds in detail, discovering that some remains represented species new to science. In 1974 Madsen named and described the type species Stokesosaurus clevelandi. Its generic name honours Stokes. The specific name refers to the town of Cleveland, Utah. The holotype (UUVP 2938) was uncovered in the Brushy Basin Member of the Morrison Formation dating from the early Tithonian stage, about 150 million years old. It consists of a left ilium or hip bone, belonging to a juvenile individual. Madsen also assigned a paratype, UUVP 2320, a 50% larger right ilium. Additionally he referred a right premaxilla, UUVP 2999. However, this was in 2005 referred to Tanycolagreus. Stokesosaurus and Tanycolagreus are about the same size, and it is possible that the latter is a junior synonym of the former. However, the ilium (the best and perhaps only known element of Stokesosaurus) of Tanycolagreus has never been recovered, making direct comparison difficult. In 1976 Peter Malcolm Galton considered Stokesosaurus to be a second species of the British possible early tyrannosauroid Iliosuchus, that he named as Iliosuchus clevelandi. This has found no acceptance among other researchers; in 1980 Galton himself withdrew his opinion. Some later finds were referred to Stokesosaurus. This included some ischia and tail vertebrae in 1991, and a partial braincase in 1998. Another, very small ilium referred to Stokesosaurus, found in South Dakota, is lost but may actually belong to the related Aviatyrannis. More fragmentary remains possibly referable to Stokesosaurus have been recovered from stratigraphic zone 2 of the Morrison Formation, dated to the late Kimmeridgian age, about 152 million years ago. A second species, Stokesosaurus langhami, was described by Roger Benson in 2008 based on a partial skeleton from England. However, further study showed that this species should be referred to a new genus, which was named Juratyrant in 2012. Benson and Stephen Brusatte concluded that not a single bone had been justifiably referred to Stokesosaurus, and that not even the paratype could be safely assigned, leaving the holotype ilium as the only known fossil of the taxon. The holotype ilium is twenty-two centimetres long, indicating a small individual. Madsen in 1974 estimated that the adult body length was about four metres. In 2010 Gregory S. Paul estimated the length at 2.5 metres, the weight at sixty kilogrammes. In 2012 Benson and Brusatte established a single unique derived trait or autapomorphy of Stokesosaurus: the normally vertical ridge on the outer blade surface of the ilium, above the hip joint, was strongly inclined to the back and reached the upper rim of the blade. In 1974 Madsen assigned Stokesosaurus to the Tyrannosauridae. However, modern cladistic analyses indicate a more basal position. In 2012 the study by Brusatte and Benson recovered Stokesosaurus as a basal member of the Tyrannosauroidea, and closely related to Eotyrannus and Juratyrant.
Размеры тела в сравнении с человеком (ювенильные особи):
Ископаемые останки (обозначены буквой B):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
| December 15th, 2012 | |
| 02:26 pm [industrialterro] [Link] |
Proceratosaurus Процератозавр (Proceratosaurus - букв. «до цератозавра») — род небольших (около 3 метров в длину) хищных ящеротазовых динозавров из семейства процератозавридов, живших в средней юре (батониан) около 167.7 - 164.7 миллионов лет назад на территории современной Англии. Изначально считался предком цератозавров (отсюда и название), в связи с аналогичным малым гребнем на его морде. Сейчас, однако, считается целурозавром, один из самых ранних известных науке, входит в надсемейство тиранозавров. Артур Смит Вудворд, который изучал процератозавра, первоначально считал его предком позднеюрского цератозавра, из-за сходства их носовых гребней. Позже исследование в 1930 году Фридриха фон Хайна позволили отнести его к группе целурозавров. До конца 1980-х годов после исследований было выявлено, что процератозавр является более примитывным тероподом, чем целурозавры, и классификацию процератозавра было решено снова пересмотреть. Грегори С. Пол предложил, что он является близким родственником орнитолеста, опять же в основном из-за гребня на носу (хотя мысль, что орнитолест рождался с носовым гребенем позже была опровергнута). Пол рассматривал процератозавра и орнитолеста, как тероподов, не относящихся ни к цератозаврам, ни к целурозаврам, и относил их к примитивным аллозаврам. Кроме того, Пол считал что динозавр Piveteausaurus относится к тому же роду, что и Proceratosaurus. Наличие на морде процератозавра маленького, как у цератозавра, рога позволило предположить, что между ними имеются родственные связи. Возможно, процератозавр питался наземными животными. Его зубы могли удерживать и скользкую рыбу. Свирепому хищнику процератозавру приходилось опасаться мегалозавра, который был в два раза крупнее и мог легко с ним справиться. Продолжительное время процератозавра относили к карнозаврам. Но, в отличие от них, его череп был мельче в тыльной верхней части. Теперь процератозавр считается ранним представителем группы целурозавров. Эта группа тероподов близких к птицам. Окаменелости динозавра были найдены около города Minchinhampton в графстве Глостершир, Англия. В длину взрослые особи процератозавра могли достигать до 3 метров, а весили около 150 килограмм. Длина черепа достигала около 30 см, что значительно меньше другого представителя семейства — килеска. Several phylogenetic studies in the early 21st century finally found Proceratosaurus (as well as Ornitholestes) to be a coelurosaur, only distantly related to the ceratosaurids and allosauroids, though one opinion published in 2000 considered Proceratosaurus a ceratosaurid without presenting supporting evidence. Phylogenetic analyses by Thomas R. Holtz, Jr. in 2004 also placed Proceratosaurus among the coelurosaurs, though with only weak support, and again found an (also weakly supported) close relationship with Ornitholestes. The first major re-evaluation of Proceratosaurus and its relationships was published in 2010 by Oliver Rauhut and colleagues. Their study concluded that Proceratosaurus was in fact a coelurosaur, and moreover a tyrannosauroid, a member of the lineage leading to the giant tyrannosaurs of the Late Cretaceous. Furthermore, they found that Proceratosaurus was most closely related to the Chinese tyrannosauroid Guanlong. They named the clade containing these two dinosaurs the Proceratosauridae, defined as all theropods closer to Proceratosaurus than to Tyrannosaurus, Allosaurus, Compsognathus, Coelurus, Ornithomimus, or Deinonychus. Этот динозавр был впервые изучен известным британским палеонтологом сэром Артуром Смитом Вудвордом в 1910 году, но он идентифицировал окаменелости как принадлежащие мегалозавру. Немецкий эксперт по динозаврам Фридрих фон Хюне столь же ошибочно приписал его к линии цератозавров, когда изучал черен пятнадцатью годами позже. Еще один экземпляр животного, которое могло быть представителем того же рода, был найден во Франции в 1923 году. Процератозавр известен только по фрагментам черепа, на котором видна основа рога на носу. Это привело фон Хюне к предположению о том, что речь идет о предке более позднего рогатого цератозара. Хотя черепной материал для изучения весьма скуден, он явно свидетельствует о том, что это карнозавр, а не цератозавр. Последние исследования показали, что процератозавр является базальным целурозавром, а также одним из древнейших предков тираннозавров.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, процератозавриды, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
| October 30th, 2012 | |
| 07:01 pm [industrialterro] [Link] |
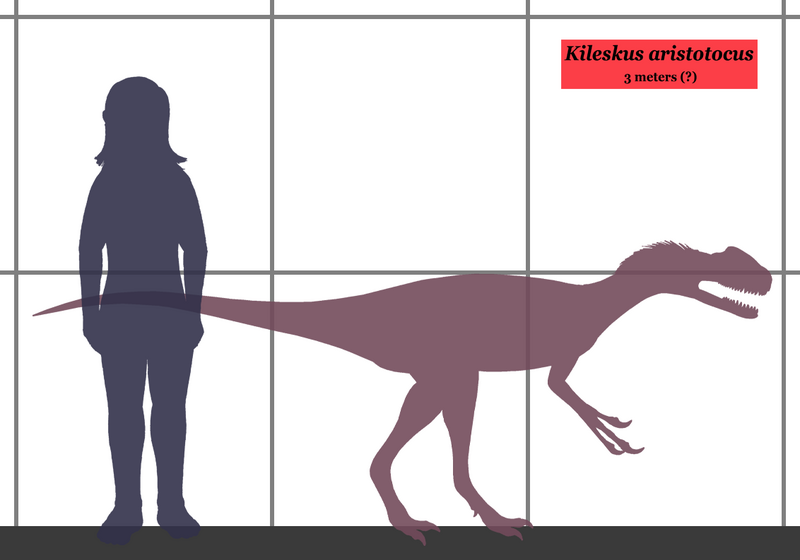
Kileskus Килеск (Kileskus; от хакасского «килескус» - ящерица) — род хищных ящеротазовых динозавров из семейства процератозавридов, живших в среднеюрском периоде (батский ярус) около 167-165 млн. лет назад на территории нынешнего Красноярского края, Россия. Типовой вид - Kileskus aristotocus. Является одним из древнейших представителей семейства, известных науке. Так же носит неофициальное название - «Шарыпозавр килескус». Искомпаемые останки были найдены в итатской свите в Березовском карьере Красноярского края, около города Шарыпово и описаны в 2010 году палеонтологами А.О. Аверьяновым, С.А. Краснолуцким и С.В. Иванцовым. Самый крупный представитель своего семейства, в длину достигал 5-6 метров. Является базальным процератозавридом, и стоит у самого основания эволюционного древа семейства. В Красноярском крае 167 миллионов лет назад жил шарыпозавр. Именно так официально назывался какое-то время единственный, найденный в России, тираннозавроид юрского периода. Палеонтологические находки в Шарыповском районе продолжают удивлять археологов. Непосредственное участие в археологических раскопках принимает местный скульптор, палеонтолог и археолог Сергей Анатольевич Краснолуцкий.
Еще в школьные годы Сергей начал увлекаться раскопками. Сейчас его находки исчисляются тысячами. Есть среди них домашняя утварь, оружие, скелеты древних животных. Один из скелетов древней мыши даже назван в его честь – мышь Краснолуцкого.
В Шарыповском краеведческом музее отведен отдельный зал под археологические и палеонтологические находки. Центральное место среди экспонатов музея занимает скелет динозавра. Узнав о том, что на берегу реки Кия томские и новосибирские ученые ведут раскопки в поисках следов древних животных, у Краснолуцкого появилась идея о том, что диназавры вполне могли водиться и в Шарыповском районе. Ведь от Шарыпово до Шестаково всего 160 км. Интуитивно Сергей Анатольевич выбрал место для поиска рядом с недавно открытым угольным разрезом. Под слоем угля он обнаружил чешую доисторических рыб, фрагменты панциря черепахи, зубы динозавров. Свои находки он отправил в Московский Палеонтологический институт для проверки их истинности.
Доктор биологических наук из Петербурга Александр Аверьянов и профессор Болонского университета Томас Мартин доказали открытие нового, ранее неизвестного вида хищных динозавров, которому вначале присвоили название шарыпозавр. Затем дали другое официальное название – kileskus aristotocus.
С точки зрения палеонтологии Шарыповский район считается самым изученным в России. Хотя финансирование из казны страны на это не выделяется. Сергей Краснолуцкий только на своем энтузиазме, затрачивая личное время и личные финансы, продолжает поиски уникальных находок. Многие из этих находок пока лежат необработанные и дожидаются того времени, когда на это будут выделены средства.
Новый вид динозавров, относящийся к надсемейству Tyrannosauroidea, описали российские палеонтологи. Окаменевшие фрагменты скелета этого динозавра – часть верхней челюсти с сохранившимся зубом, еще несколько черепных костей и кости конечностей были найдены в Березовском карьере в Красноярском крае, около города Шарыпово. Специалисты выделили килескуса в новое семейство Proceratosauridae, вместе со среднеюрским процератозавром (Proceratosaurus bradleyi), жившим на территории Великобритании и позднеюрским гуанлонгом (Guanlong wucaii) из Китая. Килескус жил в батском веке средней юры, 165-167 миллионов лет назад. Он был крупнее своих ближайших родственников, длина его черепа оценивается в 50-60 сантиметров (у процератозавра череп был всего 30 см длиной), а общая длина животного могла достигать 5-6 метров. Интересно, что килескуса можно считать древнейшим известным на данный момент "предком тираннозавра". Этот термин взят в кавычки потому что, на самом деле, нельзя утверждать, что тот или иной динозавр, живший за 100 миллионов лет до тираннозавра, был его несомненным прямым предком. Но Kileskus aristotocus является базальным процератозавридом, то есть он стоит у самого основания эволюционного древа процератозаврид, а динозавры этого семейства, в свою очередь - самые древние известные представители тираннозавроидов. Кроме того, килескус старше, чем Guanlong wucaii (тот жил в оксфордском веке, 160 миллионов лет назад) и даже, судя по всему, несколько старше чем Proceratosaurus bradleyi, живший 165 миллионов лет назад в конце батского – начале келловейского веков. Репродукции (1, 2, 3, 4, 5, 6, 7):
( Read More ) Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, процератозавриды, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
| October 22nd, 2012 | |
| 09:14 pm [industrialterro] [Link] |
Guanlong Гуанлонг (Guanlong; 五彩冠龍 — произносится как «Гуаньлун»; букв. «увенчанный короной дракон») — род хищных ящеротазовых динозавров надсемейства тираннозавровых, живших в позднем юрском периоде (около 161—159 млн лет назад) на территории нынешней провинции Синьцзян в Китае. Один из самых древних представителей надсемейства, включающий единственный вид — Guanlong wucaii. Возраст находки около 160 млн лет (Юрский период), что на 92 млн лет раньше появления более известных его родственников, таких как Tyrannosaurus. Обнаружен экспедицией палеонтологов из George Washington University в 2000 году в Китае, в провинции Синьцзян и описан в 2006 году как новый вид и род. Длина около 3 метров. Найдено два скелета. Этот двуногий теропод разделяет много общих черт с его тираннозавровыми потомками, и также имел некоторые необычные признаки, подобно большому гребню на его голове. В отличие от более поздних тиранозавров, Guanlong имел три длинных пальца на передних конечностях. Это напоминает его близкого родственника Dilong, и подобно Dilong, гуанлонги, возможно, имели оперение из примитивных перьев. Обитал на территории современной пустыни Гоби. At present, Guanlong is known from two specimens. The holotype (IVPP V14531) is a reasonably complete, partially articulated adult skeleton. Another, immature specimen is known from fully articulated and nearly complete remains. The crest on the skull of the immature specimen is notably smaller and restricted to the forward portion of the snout, while the adult has a larger and more extensive crest. The crests of both specimens are thin, delicate structures that likely served as display organs. Proceratosauridae is a family or clade of theropod dinosaurs, probably belonging to the tyrannosaur lineage. It was first named in 2010 by Oliver Rauhut and colleagues in their re-evaluation of the type genus, Proceratosaurus. Their study supported the idea that Proceratosaurus is a coelurosaur, a tyrannosauroid, and most closely related to the Chinese tyrannosauroid Guanlong. They defined the clade containing these two dinosaurs as all theropods closer to Proceratosaurus than to Tyrannosaurus, Allosaurus, Compsognathus, Coelurus, Ornithomimus, or Deinonychus. Later studies included the Russian Kileskus and the Chinese Sinotyrannus in the family. Загадочные ямы, в которых были обнаружены многочисленные кости динозавров, могут быть следами их более крупных сородичей. Три первые подобные выемки глубиной один–два метра были обнаружены около десяти лет назад в китайской провинции Синьцзян. В них находились практически полные скелеты нескольких маленьких — предков , в том числе (известен наростом на голове, напоминающим ирокез) и . Находка произвела сенсацию, так как целые скелеты тероподов встречаются очень редко (мелкие динозавры обычно попадались под ноги более крупным родичам). Но что привело к образованию подобных «братских могил»? Канадский геолог Дэвид Эберт из наконец показал, что ямы были наполнены водой и грязью из вулканического аргиллита и песчаника. «Все геологические данные свидетельствуют о том, что мы имеем дело с осадочными породами, которые первоначально были растворены в жидкости, — подчеркивает ученый. — Это были не просто дыры в почве». Когда-то пустыня Гоби, в которой и были найдены останки динозавров, представляла собой богатую водоемами и частично заболоченную местность. В конце юрского периода извержения вулканов засыпали регион огромным количеством пепла. Над водными карманами образовалась полутвердая поверхность, которую гиганты вроде пробивали без труда, и яма быстро заполнялась жидкой грязью, превращаясь в невидимую ловушку для более мелких видов. Тероподов, масса тела которых составляла всего 18–22,5 кг, застывший верхний слой вулканического мусора выдерживал без труда, но стоило животным попасть в яму, выбраться они уже не могли. Не в последнюю очередь из-за того, что для передвижения использовали только задние конечности. Четвероногие особи, скорее всего, могли спастись. Кроме того, по мнению некоторых палеонтологов, тероподы к тому времени были покрыты перьями, что еще больше снижало их шансы на выживание. Постепенно яма наполнялась погибшими животными и переставала служить ловушкой.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Read more ) Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, процератозавриды, тероподы, тираннозавроиды, целурозавры, ящеротазовые |
| August 7th, 2012 | |
| 12:41 pm [industrialterro] [Link] |
Coelurus Целюр (от Coelurus — "полый хвост") — род динозавров из средне-позднего Киммерджийского яруса Юрского периода. Хотя это название связано с группой вымерших пресмыкающихся подотряда звероногих динозавров, оно часто смешивалось с более известным Орнитолестом (Ornitholestes). Подобно многим таксонам описанным на раннем этапе развития палеонтологии, оно отличается сложой и запутанной таксономической историей. Только один вид в настоящее время считается принадлежащим этому роду, а именно типовой вид C. fragilis, описанный Отнилом Чарлзом Маршем в 1879 году по единственному неполному скелету из формации Моррисон в Вайоминге, США. Небольшой двуногий хищник с длинными ногами. Жил 153—150 млн лет назад. Целюр описан по единственному скелету, включающему многочисленные позвонки, частично сохранившиеся плечевой пояс и таз и большую часть конечностей. Скелет хранится в Музее естественной истории Пибоди. Кости были найдены в 13 карьере Рида, в Como Bluff в штате Вайоминг. Две передние конечности, предположительно от представителя этого вида, были добыты в Динозавровой шахте Кливленда Ллойда в Юте. Целюр был небольшим двуногим хищником. Предположительно, он весил от 13 до 20 килограммов и достигал в длину 2,4 метра, при высоте бедра в 70 см. Реконструкция, сделанная на основании скелета позволяет говорить о том, что целюр имел длинную шею и тело (торс), длинные и тонкие задние конечности (из-за длинной плюсны) и предположительно маленький и узкий череп. Единственной найденной частью черепа является осколок нижней челюсти, найденный там же, где и остальные кости. Хотя по степени сохранности и окраске этот осколок соответствует остальным костям, он отличается малой толщиной (длина составляет 7,9 сантиметров, при толщине в 1,1 сантиметра), что может свидетельствовать о принадлежности к другому виду. В целом, позвонки продолговатые и сплюснутые, с короткими остистыми отростками и тонкими стенками. Позвонки шейного отдела в значительной степени пневматизированы, с многочислеными выемками на поверхности. Выемки распределены неравномерно и неодинаковы по размеру. Длина позвонков шейного отдела в четыре раза превосходит ширину. Позвонки двояковогнутые. Позвонки грудного отдела менее вытянутые, не имеют выемок на поверхности, имеют менее развитые вогнутые поверхности и тела в форме песочных чаcов. Позвонки хвостового отдела также не имеют выемок на поверхности. Единственная найденная кость из плечевого пояса — лопатка. Плечо передней конечности имело S-образный изгиб и немного превосходило по длине предплечье (11.9 и 9.6 сантиметров соответственно). В запястье имеется полулунная кость, как у Deinonychus. Пальцы тонкие и длинные. Из костей таза сохранилось парные сросшиеся лобковые кости с выдающимся выростом на конце. Бедренные кости имеют S-образный изгиб при взгляде спереди (в аксиальной проекции). Кости плюсны необычно длинны и тонки, почти достигают длины бедреннной кости (длина наиболее сохранившейся бкдренной кости равна 21 сантиметру). Три наиболее известных небольших звероногих динозавров из формации Морисон в США — Целюр, Орнитолест (Ornitholestes) и Таниколагрей (Tanycolagreus) — были неспециализированными и очень сходными между собой представителями Целурозавров. В настоящее время накоплено достаточно сведений, чтобы различать их по деталям строения. Например, пропорции тела этих динозавров заметно различаются: спина и шея целюра превосходит по длине соответствующие отделы орнитолестеса. Ноги и стопы орнитолестеса короче и толще, чем у целюра. Целюр и Tanycolagreus более схожи между собой. Они различаются формой передних конечностей, предплечья и бедренных костей, расположением мест крепления скелетных мышц, длиной позвонков грудного отдела и, в особенности, длиной костей плюсны (метатарзалий). ( Далее )
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целуриды, целурозавры, ящеротазовые |
| May 30th, 2012 | |
| 08:42 pm [industrialterro] [Link] |
Aviatyrannis Aviatyrannis is a genus of carnivorous tyrannosauroid theropod dinosaur from the Kimmeridgian stage of the Late Jurassic found in Portugal. In 2000 Oliver Walter Mischa Rauhut reported the find of tyrannosauroid material in the lignite coal mine of Guimarota near Leiria, which he referred to Stokesosaurus. Later concluding the distinctiveness of the material justified a separate genus, Rauhut in 2003 named and described it as the type species Aviatyrannis jurassica. The species name was by Rauhut given the intended meaning of "tyrant's grandmother from the Jurassic". The generic name is derived from Latin avia, "grandmother", and tyrannus, "tyrant", on the presumption tyrannis would be its genitive. The specific name means "Jurassic". The holotype, IPFUB Gui Th 1, was found in a layer of the Alcobaça Formation dating from the early Kimmeridgian, about 155 million years old. It consists of a right ilium. Rauhut in 2003 referred two other bones to Aviatyrannis: IPFUB Gui Th 2, a partial right ilium, and IPFUB Gui Th 3, a right ischium. The referred elements represent slightly larger individuals. Additionally sixteen isolated teeth were referred: IPFUB GUI D 89-91: three teeth of the praemaxilla, and IPFUB GUI D 174-186: thirteen teeth of the maxilla and dentary. These had in 1998 been described by Jens Zinke. The name Aviatyrannis has not found general acceptance among Portuguese workers who often list Portuguese tyrannosauroid material of this age as a Stokesosaurus sp. On the other hand Rauhut in 2003 hypothesised that American material referred to Stokesosaurus might actually belong to Aviatyrannis. Like other early tyrannosauroids, Aviatyrannis was a rather small bipedal predator. The holotype specimen IPFUB Gui Th 1, for example, is an ilium only ninety millimeters long. It may have belonged to a juvenile. In 2010 Gregory S. Paul estimated its length at one metre, its weight at five kilogrammes. The ilium is elongated and low with the typical tyrannosauroid vertical ridge on the outer blade surface above the hip joint. The teeth of the praemaxilla have a D-shaped cross-section. The maxillary and dentary teeth are elongated, only recurving near the top, with perpendicular denticles on both edges. Their bases are circular in cross-section; the top of the tooth crown is more flattened. Aviatyrannis was in 2003 by Rauhut placed in the Tyrannosauroidea, in a basal position. Aviatyrannis is one of the oldest tyrannosauroids ever found, the oldest being Proceratosaurus (or, possibly, Iliosuchus. Целурозавры (Coelurosauria — полохвостые) — самая многочисленная группа динозавров-тероподов, живших около 167—65,5 миллионов лет назад. Раньше существовала довольно простая система классификации тероподов — деление на маленьких и больших. Маленькие тероподы назывались целурозаврами. Эта система была неправильна (эволюционные отношения не учитывались), и сейчас применяется новая классификация. По современной системе не все целурозавры маленькие — в этой группе есть и гигантские особи (например, тираннозавр и дейнохейр). Первые остатки целурозавров найдены в отложениях, датируемых средним юрским периодом. Самым ранним целурозавром был процератозавр. Другие примитивные целурозавры, орнитолестес и целур жили немного позже процератозавра. Иногда орнитолестеса, целура и процератозавра объединяют в семейство Coeluridae. Другой вариант — объединить процератозавра и орнитолестеса (оба имели своеобразный рог на голове) в семейство Ornitholestidae, целур (не имевший рога) в таком случае остаётся единственным родом в семействе Coeluridae. Вполне возможно и не объединять эти роды, тогда можно пользоваться классификацией, представленной ниже. Помимо трёх примитивных родов целурозавры включают компсогнатидов, тираннозавроидов, орнитомимозавров, манирапторов. Компсогнатиды — это мелкие, очень примитивные целурозавры. Три оставшиеся основные группы объединяются в группу тираннорапторов. Тираннорапторы делятся на тираннозавроидов и манираптороподобных (орнитомимиды + манирапторы). Тираннозавроиды — это в основном крупные хищники (хотя на начальном этапе представители группы были мелкими и покрытыми протоперьями). Орнитомимозавры — немалоизвестные, похожие на страуса, динозавры. А животные из самой развитой группы — манирапторов, в большинстве своём уже были покрыты перьями. Некоторые считают, что дромеозавридов (группа манирапторов) лучше классифицировать как птиц, а не как динозавров. Тираннозавры, или тираннозавроиды (Tyrannosauria, Tyrannosauroidea — что означает «ящерица тиран») — надсемейство хищных динозавров подотряда тероподов, живших с юрского по меловой периоды (около 167—65 млн лет назад) на территориях современных Северной Америки, Европы, Азии и Австралии. Тираннозавры были крупнейшими хищниками планеты и жили на суперконтиненте Лавразия, в начале юрского периода. К концу мелового периода, тираннозавры были доминирующими крупными хищниками в северном полушарии. Останки тираннозавров были найдены на территории Северной Америки, Европы, Азии и Австралии. Самым ранним из всех известных представителей надсемейства является — килескус, живший около 167—165 млн лет назад. С его открытием стало ясно, что не все ранние тираннозавроиды были маленькими. Длина килескуса достигала 6 метров, в то время как представитель рода Guanlong имел длину всего в 3 метра.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавроиды, целурозавры, ящеротазовые |










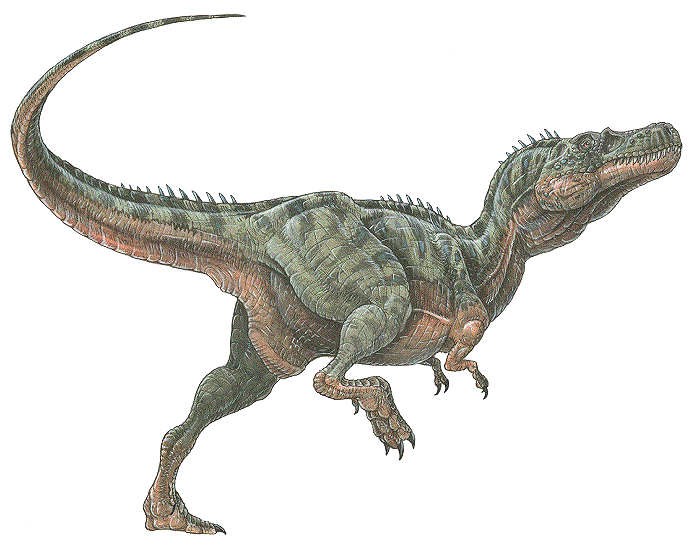




















































































{kind=link}
_from_Mongolia.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}