[Recent Entries][Archive][Friends][User Info]
| November 6th, 2012 | |
|---|---|
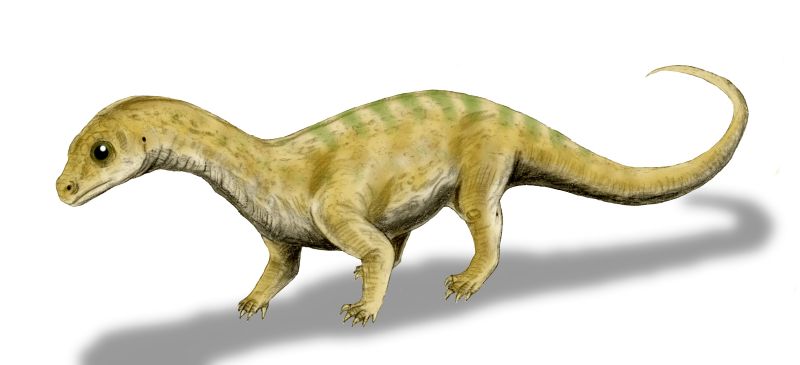
| 06:13 pm [industrialterro] [Link] | Массоспондил, или Массоспондилус (Massospondylus) — растительноядный динозавр раннего юрского периода (от 200 до 183 миллионов лет назад), один из самых древних динозавров. Название происходит от греческого μάσσων (massōn, «длинный») и σπόνδυλος (spondylos, «позвонок»). Был описан в 1854 году сэром Ричардом Оуэном по останкам, найденным в Южной Африке и является одним из первых динозавров, получивших название. Ископаемые останки были обнаружены также в Лесото, Зимбабве, Намибии, в ЮАР, США и Индии. Массоспондилы имели длинные шею и хвост, взрослые особи достигали в длину 5 метров. Во взрослом возрасте массоспондилы могли ходить на задних ногах, используя передние лапы с большим когтем на каждой для защиты и захвата листьев, употребляемых в пищу. Для добывания пищи с верхушек деревьев служила и длинная шея. Детёныши массоспондилов передвигались на четырёх ногах. Согласно современным исследованиям, после рождения они ещё не были способны добывать пищу, в связи с чем об этом должны были заботиться взрослые особи. M. kaalae Barrett, 2009 — вид, живший в юрском периоде окоо 199—189 млн лет назад. Описан в 2009 году на основе частичного черепа. Ископаемые останки были найдены в местности Voyizane в формации Upper Elliot на территории Южной Африки; M. rawesi. The type species is M. carinatus; seven other species have been named during the past 150 years, but only M. kallae among these is still considered valid. Prosauropod systematics have undergone numerous revisions during the last several years, and many scientists disagree where exactly Massospondylus lies on the dinosaur evolutionary tree. The family name Massospondylidae was once coined for the genus, but because knowledge of prosauropod relationships is in a state of flux, it is unclear which other dinosaurs—if any—belong in a natural grouping of massospondylids; several 2007 papers support the family's validity. Although Massospondylus was long depicted as quadrupedal, a 2007 study found it to be bipedal. It was probably a plant eater (herbivore), although it is speculated that the prosauropods may have been omnivorous. This animal, 4–6 meters (13–20 feet) long, had a long neck and tail, with a small head and slender body. On each of its forefeet, it bore a sharp thumb claw that was used in defense or feeding. Recent studies indicate Massospondylus grew steadily throughout its lifespan, possessed air sacs similar to those of birds, and may have cared for its young. Massospondylus was a mid-sized prosauropod that was around 4 meters (13 ft) in length and weighed approximately 135 kilograms (300 lb), although a few sources have estimated its length at up to 6 meters (20 ft). Although long assumed to have been quadrupedal, a 2007 anatomical study of the forelimbs has questioned this, arguing that their range of motion precluded effective habitual quadrupedal gait. The study also ruled out the possibility of "knuckle-walking" and other forms of locomotion that would avoid the issue of the limited ability of Massospondylus to pronate its hands. Although its mass suggests a quadrupedal nature, it would have been restricted to its hind legs for locomotion. Massospondylus was a typical prosauropod in most other respects. It possessed a slender body and long neck, with around nine long cervical (neck) vertebrae, 13 dorsal (back) vertebrae, three sacral (hip) vertebrae, and at least 40 caudal (tail) vertebrae. The pubis faced forward, as with most saurischians. It had a slighter build than that of Plateosaurus, an otherwise similar prosauropod dinosaur. A recent discovery shows that Massospondylus possessed well-developed clavicles, joined in a furcula-like arrangement, suggesting both that it had immobile shoulder blades and that clavicles were not rudimentary and nonfunctional in those dinosaurs that did not have true furculae. This discovery also indicates that the furcula of birds is derived from clavicles. Like Plateosaurus, it had five digits on each foot, with a large thumb claw used for feeding or defense against predators. The fourth and fifth digits of the forepaws were tiny, giving the forepaws a lopsided look. The 2007 study indicated that Massospondylus held its manus (hands) in a semi-supinated ("prayer-like") orientation, with the palmar surfaces facing one another; the wrist was never found rotated in articulated (still-connected) fossil specimens. The small head of Massospondylus was approximately half the length of the femur. Numerous openings, or fenestrae, in the skull reduced its weight and provided space for muscle attachment and sensory organs. These fenestrae were present in pairs, one on each side of the skull. At the front of the skull were two large, elliptical nares. The orbits were proportionally larger in Massospondylus than in related genera such as Plateosaurus. The antorbital fenestrae, smaller than those seen in Plateosaurus, were situated between the eyes and the nose. At the rear of the skull were two more pairs of temporal fenestrae: the lateral temporal fenestrae immediately behind the eye sockets and the supratemporal fenestrae on top of the skull. Small fenestrae also penetrated each mandible. The shape of the skull is traditionally restored as wider and shorter than that of Plateosaurus, but this appearance may be due just to differential crushing experienced by the various specimens. Some features of the skull are variable between individuals; for example, the thickness of the upper border of the orbit and the height of the posterior maxilla. These differences may be due to sexual dimorphism or individual variation. As with other prosauropods, it has been proposed that Massospondylus had cheeks. This theory was proposed because there are a few large holes for blood vessels on the surfaces of the jaw bones, unlike the numerous small holes present on the jaws of cheekless reptiles. The cheeks would have prevented food from spilling out when Massospondylus ate. Crompton and Attridge (1986) described skulls of Massospondylus as possessing pronounced overbites and suggested the presence of a horny beak on the tip of the lower jaw to make up the difference in length and account for tooth wear on the teeth at the tip of the snout. However, this was later shown to be a misinterpretation based on crushing in a top–bottom plane. Skulls not crushed in this orientation do not show an overbite. There also seems to be some variation of tooth morphology based on the position of teeth in the jaw. The heterodonty present in Massospondylus is greater than that present in Plateosaurus, although unsurprisingly not as pronounced as the specialization of teeth in Heterodontosaurus. Teeth closer to the front of the snout had round cross-sections and tapered to points, unlike the back teeth, which were spatulate and had oval cross-sections. Massospondylus is a prosauropod, a grouping of early saurischian dinosaurs that lived during the Triassic and Jurassic, but which had died out by the end of the Jurassic. Other members of the group include Plateosaurus, Yunnanosaurus, and Riojasaurus. Basal sauropodomorph systematics continue to undergo revision, and many genera once considered classic prosauropods have recently been removed from the group in phylogenetic nomenclature, on the grounds that their inclusion would not constitute a clade (a natural grouping containing all descendants of a single common ancestor). Exactly which animals constitute a monophyletic (natural grouping) of prosauropods is unclear. Yates and Kitching (2003) published a clade consisting of Riojasaurus, Plateosaurus, Coloradisaurus, Massospondylus, and Lufengosaurus. Galton and Upchurch (2004) included Ammosaurus, Anchisaurus, Azendohsaurus, Camelotia, Coloradisaurus, Euskelosaurus, Jingshanosaurus, Lessemsaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Mussaurus, Plateosaurus, Riojasaurus, Ruehleia, Saturnalia, Sellosaurus, Thecodontosaurus, Yimenosaurus and Yunnanosaurus in a monophyletic Prosauropoda. Wilson (2005) considered Massospondylus, Jingshanosaurus, Plateosaurus, and Lufengosaurus a natural group, with Blikanasaurus and Antetonitrus possible sauropods. Bonnan and Yates (2007) considered Camelotia, Blikanasaurus and Melanorosaurus possible sauropods. Yates (2007) placed Antetonitrus, Melanorosaurus, and Blikanasaurus as basal sauropods and declined to use the term Prosauropoda, as he considered it synonymous with Plateosauridae. However, he did not rule out the possibility that a small group of prosauropods consisting of Plateosaurus, Riojasaurus, Massospondylus and their closest kin were monophyletic. Massospondylus is the type genus of the proposed family Massospondylidae, to which it gives its name. The Massospondylidae family may also include Yunnanosaurus, although Lu et al. (2007) placed Yunnanosaurus in its own family. Yates (2007) considered Massospondylus, Coloradisaurus, and Lufengosaurus massospondylids, with Yunnanosaurus in Anchisauria. Smith and Pol (2007) also found a Massospondylidae in their phylogenetic analysis, including Massospondylus, Coloradisaurus, and Lufengosaurus, as well as their new genus, Glacialisaurus. Adeopapposaurus, based on the fossils once thought to belong to a South American form of Massospondylus, was also classified as a massospondylid. Pradhania was originally regarded as a basal sauropodomorph but new cladistic analysis performed by Novas et al., 2011 suggests that Pradhania is a massospondylid. Pradhania presents two synapomorphies of Masospondylidae recovered in their phylogenetic analysis and its fossils were discovered from the same region and basin in India as M. hislopi. The faunas and floras of the Early Jurassic were similar worldwide, with conifers adapted for hot weather becoming the common plants, and prosauropods and basal theropods the main constituents of a worldwide dinosaur fauna. The environment of early Jurassic southern Africa has been described as a desert. African Massospondylus was a contemporary of early crocodylomorphs; tritylodontid and trithelodontid therapsids; morganucodontid mammals; and dinosaurs including the small theropod Megapnosaurus rhodesiensis; a species of Melanorosaurus (M. thabanensis); and several genera of early ornithischians, such as Lesothosaurus and the heterodontosaurids Abrictosaurus, Heterodontosaurus, and Lycorhinus. It is not clear which carnivores may have preyed on Massospondylus. Most of the theropods which have been discovered in rocks of Early Jurassic age in southern Africa, such as Megapnosaurus, were smaller than mid-sized prosauropods like Massospondylus. These smaller predators have been postulated as using fast slashing attacks to wear down prosauropods, which could have defended themselves with their large hand and foot claws. The 6-meter-(20 ft-) long carnivorous theropod Dracovenator lived during the same period (Hettangian to Sinemurian stages) as Massospondylus and has also been found in the Elliot Formation of South Africa. As with all dinosaurs, much of the biology of Massospondylus, including its behavior, coloration, and physiology, remains unknown. However, recent studies have allowed for informed speculation on subjects such as growth patterns, diet, posture, reproduction, and respiration. A 2007 study suggested that Massospondylus may have used its short arms in defense from predators ("defensive swats"), in intraspecies combat, or in feeding, although its arms were too short to reach its mouth. Scientists speculate that Massospondylus could have used its large pollex (thumb) claw in combat, to strip plant material from trees, digging, or for grooming. А 2005 study indicated that Massospondylus' sister taxon, Plateosaurus, exhibited growth patterns affected by environmental factors. The study indicated that when food was plentiful, or when the climate was favorable, Plateosaurus exhibited accelerated growth. This pattern of growth is called "developmental plasticity". It is unseen in other dinosaurs, including Massospondylus, despite the close relationship between Plateosaurus and Massospondylus. The study indicated that Massospondylus grew along a specific growth trajectory with little variation in the growth rate and ultimate size of an individual. Another study of age determination indicated that Massospondylus grew at a maximum rate of 34.6 kg (76.3 lb) per year and was still growing at around 15 years of age. Prosauropods such as Massospondylus may have been herbivorous or omnivorous. As recently as the 1980s, paleontologists debated the possibility of carnivory in prosauropods. However, the hypothesis of carnivorous prosauropods has been discredited, and all recent studies favor a herbivorous or omnivorous lifestyle for these animals. Galton and Upchurch (2004) found that cranial characteristics (such as jaw articulation) of most prosauropods are closer to those of herbivorous reptiles than those of carnivorous ones, and the shape of the tooth crown is similar to those of modern herbivorous or omnivorous iguanas. The maximum width of the crown was greater than that of the root, resulting in a cutting edge similar to those of extant herbivorous or omnivorous reptiles. Barrett (2000) proposed that prosauropods supplemented their herbivorous diets with small prey or carrion. Gastroliths (gizzard stones) have been found in association with Massospondylus fossils in South Africa, and with a Massospondylus-like animal from the Late Triassic of Virginia. Massospondylus swallowed these stones to aid in digestion; muscular contractions in the creature's gizzard would have pulverized swallowed plant material, compensating for its inability to chew. In 1977, seven 190-million-year-old Massospondylus eggs were found in Golden Gate Highlands National Park in South Africa by James Kitching, who identified them as most likely belonging to Massospondylus. It was nearly 30 years before extraction was started on the fossils of the 15-centimeter- (6 in-) long embryos. They remain the oldest dinosaur embryos ever found. By early 2012, at least 10 nests from at least four fossiliferous horizons had been found, with up to 34 eggs per nest. Sedimentary structures indicate that the nesting area was in a shoreline setting. Notably, the near-hatchlings had no teeth, suggesting they had no way of feeding themselves. Based on the lack of teeth and the animal's body proportions, scientists speculate that postnatal care might have been necessary. This is supported by evidence that the hatchlings remained at the nest sites until they had doubled in size. The four legs of the near-hatchlings were of equal length, indicating that newly hatched Massospondylus were quadrupedal. The discovery of hatchling footprints confirmed their quadrupedality. The skull and eyes were proportionately oversized when compared to adults, as is common in other very young vertebrates, although the skulls of juveniles were taller and more narrow as well. The quadrupedality of the hatchlings suggests that the quadrupedal posture of later sauropods may have evolved from retention of juvenile characteristics in adult animals, an evolutionary phenomenon known as pedomorphosis. The nesting sites may have been used repeatedly (site fidelity), by groups of animals (colonial nesting); in both cases, these represent the oldest evidence of the behaviors. Many saurischian dinosaurs possessed vertebrae and ribs that contained hollowed-out cavities (pneumatic foramina), which reduced the weight of the bones and may have served as a basic 'flow-through ventilation' system similar to that of modern birds. In such a system, the neck vertebrae and ribs are hollowed out by the cervical air sac; the upper back vertebrae, by the lung; and the lower back and sacral (hip) vertebrae, by the abdominal air sac. These organs constitute a complex and very efficient method of respiration. Prosauropods are the only major group of saurischians without an extensive system of pneumatic foramina. Although possible pneumatic indentations have been found in Plateosaurus and Thecodontosaurus, the indentations were very small. One study in 2007 concluded that prosauropods like Massospondylus likely had abdominal and cervical air sacs, based on the evidence for them in sister taxa (theropods and sauropods). The study concluded that it was impossible to determine whether prosauropods had a bird-like flow-through lung, but that the air sacs were almost certainly present.
Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9, 10):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, массоспондилиды, прозавроподы, ящеротазовые |















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
,_Golden_Gate_National_Park,_South_Africa,_Early_Jurassic_-_Royal_Ontario_Museum_-_DSC00145.JPG){kind=link}