[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 20 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| October 25th, 2012 | |
|---|---|
| 07:16 pm [industrialterro] [Link] |
Jobaria Джобария (Jobaria) — род вымерших ящеротазовых динозавров из группы зауропод-макронарий (прим. - на самом деле, неясного систематического положения), живших в юрском периоде (около 175,6—145,5 миллионов лет назад), на территории нынешней Африки. Окаменелости зауропода были найдены в пустыне Сахара в местности Agadez (формация Tiouraré), Нигер. Впервые описан палеонтологом Полом Серено в 1999 году. Представлен одним видом — Jobaria tiguidensis. Джобария была крупным динозавром: Длина тела — 21 метр, высота — 6 метров, вес — 20 тонн. Внешне джобария была похожа на камаразавра. Толстое тело, длный хвост, толстые ноги, шея была не очень длинной, а голова была небольшой. Jobaria was a genus of sauropod dinosaur that lived in the current day Niger during the middle Jurassic Period, between 164-161 million years ago. Jobaria was a primitive sauropod, around 21.3 metres (70 ft) long and estimated to weight about 22.4 tonnes (25 short tons). Its backbone and tail were simple compared to the complex vertebrae and whiplash tail of the later North America sauropods Diplodocus and Apatosaurus. It may also been able to rear up on its hind legs as Paul Sereno concluded, after comparing the ratios of humerus and femur circumferences in Jobaria to extant elephants. The weight distribution of Jobaria indicates that it was supported by the rear limbs rather than the forelimbs (as in elephants) and is speculated that as elephants can rear up, then Jobaria would have been able to more easily. The phylogenetic relationships of Jobaria are uncertain, it has been interpreted either as a basal macronarian, or as a non-neosauropod eusauropod, basal to the neosauropod clade.
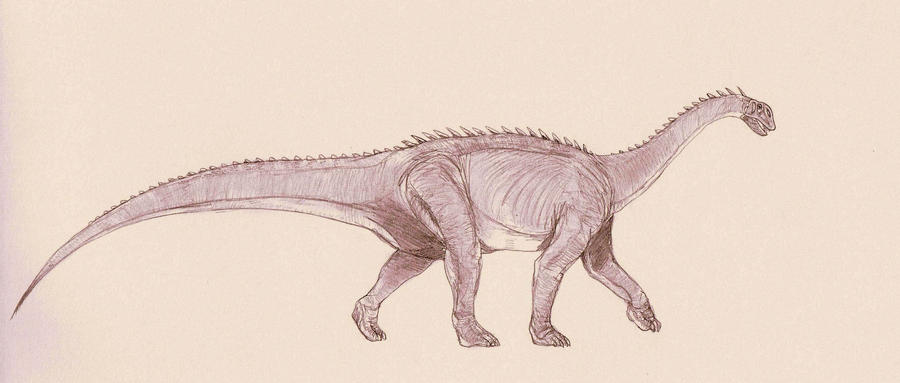
Стадо джобарий отбивается от афровенаторов
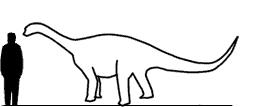
Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| October 23rd, 2012 | |
| 07:30 pm [industrialterro] [Link] |
Haplocanthosaurus Haplocanthosaurus (meaning "simple spined lizard") is a genus of sauropod dinosaur. Two species, H. delfsi and H. priscus, are known from incomplete fossil skeletons. It lived during the late Jurassic period (Kimmeridgian stage), 155 to 152 million years ago. The type species is H. priscus, and the referred species H. delfsi was discovered by a young college student named Edwin Delfs in Colorado. Haplocanthosaurus specimens have been found in the very lowest layer of the Morrison Formation, along with Hesperosaurus, Eobrontosaurus, and Allosaurus jimmadensi. Haplocanthosaurus was one of the smallest sauropods of the Morrison. While some Morrison sauropods could reach lengths of over 20 meters (or over 70 feet), Haplocanthosaurus wasn't nearly as large, and reached a total length of 14.8 meters (49 feet) and an estimated weight of 12.8 metric tons. There are four known specimens of Haplocanthosaurus: one of H. delfsi, and three of H. priscus. Of these, the type of H. delfsi is the only one complete enough to mount. The mounted specimen of H. delfsi now stands in the Cleveland Museum of Natural History, albeit with a completely speculative replica skull, as the actual skull was not recovered. Present in stratigraphic zones 1, 2, and 4. Haplocanthosaurus priscus was originally named Haplocanthus priscus by John Bell Hatcher in 1903. Soon after his original description, Hatcher came to believe the name Haplocanthus had already been used for a genus of acanthodian fish (Haplacanthus, named by Louis Agassiz in 1845), and was thus preoccupied. Hatcher re-classified his sauropod later in 1903, giving it the new name Haplocanthosaurus. However, the name was not technically preoccupied at all, since there was a variation in spelling: the fish was named Haplacanthus, not Haplocanthus. While Haplocanthus technically remained the valid name for this dinosaur, Hatcher's mistake was not noticed until many years after the name Haplocanthosaurus had become fixed in scientific literature. When the mistake was finally discovered, a petition was sent to the ICZN (the body which governs scientific names in zoology), which officially discarded the name Haplocanthus and declared Haplocanthosaurus the official name (ICZN Opinion #1633). Originally described as a "cetiosaurid", José Bonaparte decided in 1999 that Haplocanthosaurus differed enough from other sauropods to warrant its own family, the Haplocanthosauridae. Phylogenetic studies have failed to clarify the exact relationships of Haplocanthosaurus with any certainty. Studies have variously found it to be more primitive than the neosauropods, a primitive macronarian (related to the ancestor of more advanced forms such as Camarasaurus and the brachiosaurids), or a very primitive diplodocoid, more closely related to Diplodocus than to titanosaurs, but more primitive than rebachisaurids.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |
| October 22nd, 2012 | |
| 08:43 pm [industrialterro] [Link] |
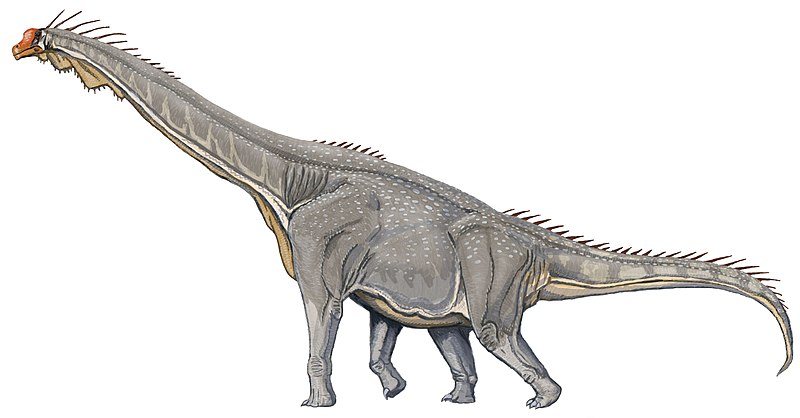
Giraffatitan Жираффатитан (Giraffatitan brancai) — гигантский динозавр-зауропод из семейства брахиозавров. Описан в 1914 году Яненшем под названием Brachiosaurus brancai из позднеюрских (киммеридж) отложений Тендагуру на территории нынешней Танзании. Скелет был смонтирован в Берлинском музее Естественной истории и является самым высоким смонтированным скелетом динозавра. В отдельный род выделен Дж. Ольшевским в 1991 г., но это выделение не получило широкого признания и чаще новый род рассматривается в качестве синонима рода Brachiosaurus. Выделение рода жираффатитан обусловлено отличиями в строении черепа и скелета. От брахиозавра отличается также относительно лёгким телосложением. При длине около 25 метров весил лишь 15—30 тонн. Высота могла превышать 15 метров, но по современным данным, брахиозавры не могли держать шею вертикально. Таким образом, реальная высота головы над землёй была меньше. Высота в плечах у крупных особей составляла около 7 метров. В отличие от американского брахиозавра, известен по многочисленным скелетам разного размера (вёл стадный образ жизни). Характерен для нижних и средних слоёв отложений Тендагуру. Вероятно, питался листвой деревьев. Гребень на черепе выше, чем у американского вида, мог служить для терморегуляции или вокализации (на гребне могли быть закреплены кожистые голосовые мешки). Вряд ли умел хорошо плавать (как показано на старых реконструкциях) ввиду неуклюжести жирафоподобного телосложения и узких подошв стоп и кистей. В целом известен лучше, чем американский вид, часто изображается под названием «брахиозавр». Giraffatitan was a sauropod, one of a group of four-legged, plant-eating dinosaurs with long necks and tails and relatively small brains. It had a giraffe-like build, with long forelimbs and a very long neck. The skull had a tall arch anterior to the eyes, consisting of the bony nares, a number of other openings, and "spatulate" teeth (resembling chisels). The first toe on its front foot and the first three toes on its hind feet were clawed. Traditionally, the distinctive high-crested skull has been seen as a characteristic of the genus Brachiosaurus to which Giraffatitan brancai was originally referred, but because within the traditional Brachiosaurus material it is known only from Tanzanian specimens now assigned to Giraffatitan, it is possible that Brachiosaurus altithorax did not show this feature. For many decades, Giraffatitan was claimed to be the largest dinosaur known, (ignoring the possibly larger but lost Amphicoelias) but in the later part of the 20th century several giant titanosaurians (Argentinosaurus, Puertasaurus and Futalognkosaurus) appear to have surpassed Giraffatitan in terms of sheer mass. However, Giraffatitan, along with Brachiosaurus are still the largest dinosaurs known from relatively complete material. All size estimates for Giraffatitan are based on the specimen HMN SII, a subadult individual between 21.8–22.5 metres (72–74 ft) in length. Mass estimates are more problematic and historically have varied from as little as 15 tonnes (17 short tons) to as much as 78 tonnes (86 short tons). These extreme estimates are now considered unlikely due to flawed methodologies. More recent estimates based on models reconstructed from bone volume measurements, which take into account the extensive, weight-reducing air sac systems present in sauropods, and estimated muscle mass, are in the range of 23–37 tonnes (25–41 short tons). However, HMN SII is not the largest specimen known (an assertion suported by its subadult status) but HMN XV2, represented by a tibia 13% larger than the corresponding material on HMN SII, which might have attained 26 metres (85 ft) in length. Like other sauropods, Giraffatitan had a relatively small brain, when its massive body size is taken into account, of about 300 cm³. A 2009 study calculated its brain-to-body mass ratio (a rough estimate of possible intelligence) at a low 0.62 or 0.79, depending on the size estimate used. Giraffatitan is also similar to other sauropods in having an enlargement of the spinal cord above the hips, which some older sources misleadingly referred to as a "second brain". The placement of Giraffatitan nostrils has been the source of much debate with Witmer (2001) describing in Science the hypothesized position of the fleshy nostrils in Giraffatitan in as many as five possible locations. There has also been the hypothesis of various sauropods, such as Giraffatitan, possessing a trunk. The fact that there were no narrow-snouted sauropods (Giraffatitan included) tends to discredit such a hypothesis. Stronger evidence for the absence of a trunk is found in the teeth wear of Giraffatitan, which shows the kind of wear that would result from biting and tearing off of plant matter rather than purely grinding, which would be the result of having already ripped the leaves and branches off with its trunk. If Giraffatitan was endothermic (warm-blooded), it would have taken an estimated ten years to reach full size, if it were instead poikilothermic (cold-blooded), then it would have required over 100 years to reach full size. As a warm-blooded animal, the daily energy demands of Giraffatitan would have been enormous; it would probably have needed to eat more than ~182 kg (400 lb) of food per day. If Giraffatitan was fully cold-blooded or was a passive bulk endotherm, it would have needed far less food to meet its daily energy needs. Some scientists have proposed that large dinosaurs like Giraffatitan were gigantotherms. The nostrils of Giraffatitan, like the huge corresponding nasal openings in its skull, were long thought to be located on the top of the head. In past decades, scientists theorized that the animal used its nostrils like a snorkel, spending most of its time submerged in water in order to support its great mass. The current consensus view, however, is that Giraffatitan was a fully terrestrial animal. Studies have demonstrated that water pressure would have prevented the animal from breathing effectively while submerged and that its feet were too narrow for efficient aquatic use. Furthermore, new studies by Lawrence Witmer (2001) show that, while the nasal openings in the skull were placed high above the eyes, the nostrils would still have been close to the tip of the snout (a study which also lends support to the idea that the tall "crests" of brachiosaurs supported some sort of fleshy resonating chamber).
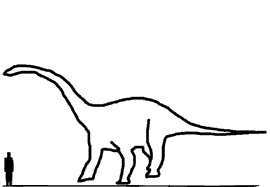
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, брахиозавриды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| October 18th, 2012 | |
| 07:18 pm [industrialterro] [Link] |
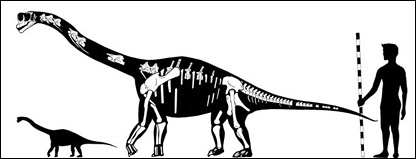
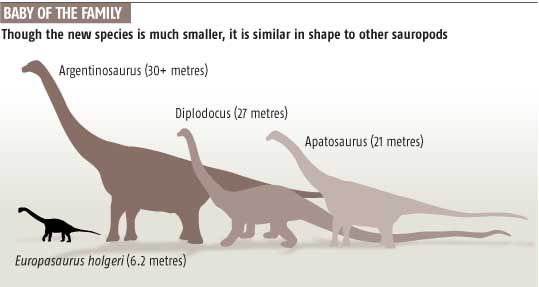
Europasaurus Europasaurus is a basal macronarian sauropod, a form of quadrupedal herbivorous dinosaur. It lived during the Late Jurassic (Kimmeridgian, between 155.7 and 150.8 million years ago) of northern Germany, and has been identified as an example of insular dwarfism resulting from the isolation of a sauropod population on an island within the Lower Saxony basin. Europasaurus holgeri exhibits the following unambiguous autapomorphic states: "nasal process of premaxillary projecting anterodorsally; medial notch is posterior dorsal margin of cervical vertebral centra; scapular acromion with a prominent posterior projection; and transverse width of astragalus twice its dorsoventral height and anteroposterior width." In comparing Europasaurus with Camarasaurus, Europasaurus differs "in the wing-shaped posterior process of the postorbital being slightly longer and wider than the anterior process, whereas it is much shorter in Camarasaurus." Other differences include Europasaurus' shorter nasal-frontal contact and a parietal bone which is rectangular in posterior view. Unlike Camarasaurus, the neural spines of the presacral vertebrae are undivided. In comparing Europasaurus with Brachiosaurus, the former genus differs from the latter by having a shorter muzzle, a quadratojugal which makes contact with the squamosal, and an anteromedially flattened humerus possessing unaligned proximal and distal epiphyses. Europasaurus was also compared with the macronarians Lusotitan and "Cetiosaurus" humerocristatus and found to be distinct from both those forms. Finally, Europasaurus can be distinguished from most known members of the Neosauropoda by its very small adult body size. Remains of both adults and juveniles have been recovered from marine carbonate beds ("bed 93"), representing more than eleven individuals ranging from 1.7 to 6.2 meters in total body length. The genus name means "lizard from Europe" (Europe + Greek sauros = lizard), and the species is named in honor of Holger Lüdtke, who discovered the first fossils of the taxa. The holotype specimen (DFMMh/FV 291; Dinosaurier-Freilichtmuseum Münchehagen/Verein zur Förderung der Niedersächsischen Paläontologie) consists of portions of a disarticulated skull, along with cervical and sacral vertebrae, derived from a single individual. This specimen and all others referred to the taxon were collected from "bed 93" at the Langenberg quarry, Oker near Goslar, Lower Saxony (Niedersachsen). A cladistic analysis of Europasaurus holgeri indicates that the species' dwarfism is a derived characteristic, and that this taxon is a more derived macronarian than Camarasaurus, and should be regarded as the sister group of the Brachiosauridae and all more derived members of the Titanosauriformes. It has been suggested that the ancestor of Europasaurus would have "dwarfed rapidly" after immigrating to a paleo-island, as even the largest of these islands in the Lower Saxony basin (a landmass <2,000 square kilometers) would not have possessed sufficient food for a population of large sauropods. Other examples of insular dwarfism among dinosaurs have been cited from the Maastrichtian (latest Cretaceous) Haţeg Island in Romania, namely the sauropod Magyarosaurus and the hadrosaur Telmatosaurus. In these two examples, paleogeography and paleoecology have been employed to explain the evolution of new and miniature versions of larger species. These fossils were determined to represent a dwarf species, and not merely juveniles of a large macronarian species, by studying the histology of the fossil bones. Likewise, by comparing the long-bone histology of a typical large-bodied sauropod, Camarasaurus, paleontologists have concluded that the diminutive size of Europasaurus resulted from a decreased growth rate. This phenomenon is a reversal of the accelerated growth responsible for gigantism in other sauropod dinosaurs. Dwarfism among macronarians seems especially remarkable as this clade includes some of the largest known dinosaurs, such as the brachiosaurs Brachiosaurus and Sauroposeidon.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11):
( Далее ) Размеры тела в сравнении с человеком (впереди неполовозрелая особь):
Размеры тела в сравнении с некоторыми другими завроподами (закрашен чёрным):
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |
| October 10th, 2012 | |
| 08:24 pm [industrialterro] [Link] |
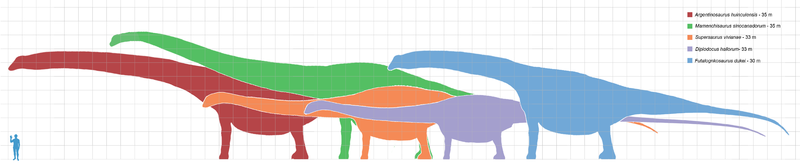
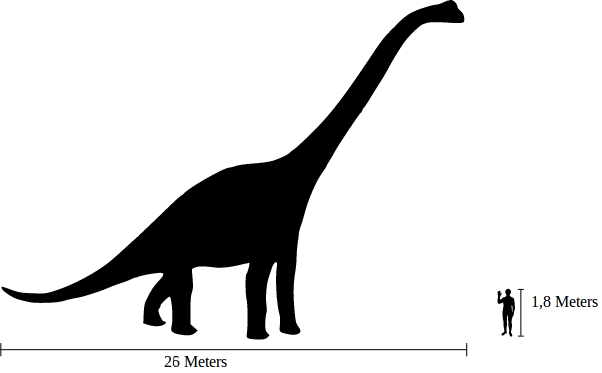
Diplodocus Диплодо́к (Diplodocus) — род ящеротазовых динозавров из группы зауропод. Первый окаменелый скелет был найден в 1877 году в Скалистых горах (Колорадо) палеонтологом С. У. Уилистоном (Samuel Wendell Williston). Позже были обнаружены и другие останки в отложениях юрского периода (формация Моррисон). Все они датируются возрастом 150—147 млн лет назад. Является крупнейшим из динозавров, известных по полным скелетам. Название рода происходит от греческих слов διπλόος — «сдвоенная» и δοκός — «Балка», дано из-за двойных нижних остистых отростков позвонков. Один из самых больших динозавров позднеюрского периода. Диплодок достигал в длину 27 метров, но по мнению ученых размеры самых крупных особей могли и вовсе достигать 35 метров. Из них большая часть приходилась на шею и хвост. Кости шеи и хвоста у диплодока были полыми. Вес диплодока по одним оценкам составлял 10—20 тонн, а по другим достигал 20—80 тонн. Диплодок имел длинную шею, состоящую из 15 позвонков, возможно, заполненных сообщающимися воздушными мешками. Череп диплодока имел непарное носовое отверстие, расположенное не на кончике морды, а в верхней части головы впереди глаз. Зубы в форме узких лопаточек имелись только в передней части рта. Конечности диплодока были пятипалыми, с короткими массивными когтями на внутренних пальцах. Длинный хвост диплодока, заканчивавшийся тонким «хлыстом», служил прекрасным орудием защиты. Вероятно, диплодоки вели стадный образ жизни, питаясь листьями невысоких деревьев. Не умея жевать, они заглатывали камни, которые помогали им перетирать пищу. Подобно брахиозавру, диплодок передвигался на четырех ногах, причём задние были длиннее передних. За период с 1878 по 1924 годы было описано несколько видов, относящихся к роду Диплодок. Первый скелет обнаружили Benjamin Mudge и Samuel Wendell Williston в 1878 году на западе США, в штате Вайоминг. По этому экземпляру известный палеонтолог того времени Г. Марш описал новый вид, назвав его Diplodocus longus. Впоследствии фоссилии диплодоков были найдены и на территории других западных штатов: Колорадо, Юта и Монтана. В музее естественной истории Карнеги находится череп юного диплодока. Этот маленький череп был обнаружен в 1921 году. Исследовав его, специалисты по палеонтологии в 2010 году пришли к выводу, что форма головы диплодока сильно менялась по мере роста. Известно несколько видов диплодоков, все виды — растительноядные формы. В современных классификациях к диплодокам относят также сейсмозавра (Diplodocus hallorum). ( Далее ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12):
( Далее ) Размеры тела в сравнении с другими гигантскими завроподами и человеком (закрашен зелёным цветом):
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, диплодоциды, завроподоморфы, завроподы, ящеротазовые |
| October 9th, 2012 | |
| 08:03 pm [industrialterro] [Link] |
Dinheirosaurus Dinheirosaurus (meaning "Porto Dinheiro lizard") is a genus of diplodocid sauropod dinosaur. Its remains were found in the upper section (Kimmeridgian) of the Late Jurassic strata of the Camadas de Alcobaça Formation located in central-western Portugal at Porto Dinheiro, Lourinhã. The fossils were described in 1999 by Bonaparte and Mateus, after excavations that lasted from 1987, the year of its discovery, to 1992. The material found was at first referred to the previously described and coeval genus Lourinhasaurus due to perceived similarities. Later analysis revealed the fossil to be a different kind of sauropod and was thus described as Dinheirosaurus. The two genera were for some time still regarded as possible synonyms but further interpretation of both finds left no doubt of their independent status. The type species is D. lourinhanensis, named for the town of Lourinhã.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, диплодоциды, завроподоморфы, завроподы, ящеротазовые |
| August 9th, 2012 | |
| 07:00 pm [industrialterro] [Link] |
Dicraeosaurus Дикреозавр (Dicraeosaurus) — растительноядный динозавр позднеюрской эпохи, представитель завропод. Близкий родственник диплодока, относится к семейству Dicraeosauridae в подотряде Diplodocoidea. Мелкий завропод, длиной до 13 метров, весом до 3—4 тонн. Отличается относительно короткой шеей и крупной головой. Характерны очень высокие раздвоенные на вершине остистые отростки. Передние конечности небольшие, но мощные. Один из наиболее примитивных известных представителей группы диплодоков. Тем не менее, дикреозавры характерны для позднеюрских (киммеридж) отложений — они представляют собой выжившую группу примитивных диплодоков. Собственно дикреозавр известен только из поздней юры Танзании (Тендагуру). Типовой вид — D. hansemanni, характерен для средних слоев Тендагуру, составляет примерно 22 % всех находок завропод в этих отложениях. Вероятно, обитали большими стадами, питались низкой растительностью. Второй вид D. sattleri — несколько более молодой по возрасту, относящийся к верхним слоям Тендагуру. Это очень мелкий завропод, не более 1 тонны весом. У него еще выше остистые отростки в области шеи. Этот вид является переходным к другому представителю дикреозавров — амаргазавру (ранний мел Аргентины). У амаргазавра (Amargasaurus cazaui) остистые отростки на шее образуют высокие вилкообразные шипы. Возможно, шипы поддерживали своеобразный «парус» или служили для защиты. Амаргазавр не превышал 10—12 метров в длину. Другие дикреозавры — брахитрахелопан (Brachytrachelopan mesai) из позднеюрских титонских отложений Аргентины и амазоназавр (Amazonsaurus marahensis) из раннего мела Бразилии. Брахитрахелопан — завропод с самой короткой шеей. Это мелкое животное своей шеей напоминало скорее гадрозавра, чем завропода. Таким образом, дикреозавры представляют гондванскую группу диплодоков. Их существование ограничено поздней юрой — ранним мелом. Ранее к этой группе относили также некоторых завропод из позднего мела Монголии (немегтозавр), но сейчас существует также точка зрения, согласно которой они принадлежат к титанозаврам. Dicraeosaurus lived in the Late Jurassic. It was herbivorous; however, it didn’t compete with other dinosaurs for vegetation. Fossils have been discovered in the rocks of Tendaguru Hill in Tanzania. The rocks also yield fossils of Giraffatitan and Kentrosaurus. As there was a distinct difference in size between these animals, they would probably have browsed for vegetation at different levels, allowing them to co-exist without significant competition Близкий родственник диплодока, относится к семейству Dicraeosauridae в подотряде Diplodocoidea. Мелкий завропод, длиной до 13 метров, весом до 3-4 тонн. Отличается относительно короткой шеей и крупной головой. Характерны очень высокие раздвоенные на вершине остистые отростки позвонков. Передние конечности небольшие, но мощные. По сравнению с поздними формами завроподов, дикреозавр имел короткие шею и хвост, и относительно крупную голову. В его позвонках также можно отметить несколько особенностей, например, имелись необычные, Y-образные шипы по всей длине спины и на шее. Возможно, они образовывали ясно видный гребень. Они использовались, чтобы быть опорой для мощных мускулов шеи и хвоста. Шея была очень коротка, она состояла только из 11 или 12 позвонков, а голова была необычно большая по отношению к телу. Зубы сидели по краям челюстей и были похожими на короткие карандаши, приспособленные для срезания растительности. Репродукции (1, 2, 3, 4, 5, 6, 7):
Размеры тела в сравнении с человеком и другими дикреозаврами:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, дикреозавры, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| 06:50 pm [industrialterro] [Link] |
Datousaurus Datousaurus, meaning either "Chieftain lizard" or "Big-head Lizard" (from the Malay datou "chieftain" or Chinese da tou "Big Head" and Greek sauros/σαυρος "lizard") was a dinosaur from the Middle Jurassic. It was a sauropod collected from the Lower Shaximiao Formation in Dashanpu, Zigong Sichuan province, China. It shared the local Middle Jurassic landscape with other sauropods Shunosaurus, Omeisaurus and Protognathosaurus, the ornithopod Xiaosaurus and the early stegosaur Huayangosaurus as well as the carnivorous Gasosaurus. Datousaurus was named by Dong Zhiming and Tang in 1984. To date, only two partial skeletons have been discovered. Neither had an articulated skull, although one skull has been discovered that has been attributed to the genus. D. bashanensis is the only established species. Datousaurus was about 15 metres long and herbivorous. It had a deep large skull for a sauropod. The rarity of its fossils suggest that it may not have been as social as other sauropods, which are often preserved in large numbers in a single deposit. Datousaurus and Shunosaurus were both closely related animals with similar anatomies. However, Datousaurus's elongated vertabrae gave it a higher reach and its teeth were more spoon shaped. This may be a sign that these contemporaries fed on different plants and/or at different heights in the trees. This strategy may have reduced competition between the two genera. A similar pattern of height difference possibly associated with feeding behaviors is found in the diplodocids.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, маменчизавриды, ящеротазовые |
| August 6th, 2012 | |
| 12:43 pm [industrialterro] [Link] |
Chuanjiesaurus Chuanjiesaurus is a genus of sauropod dinosaurs from the middle Jurassic Period. They lived in what is now China. The type species, Chuanjiesaurus ananensis, was first described by Fang, Pang, Lü, Zhang, Pan, Wang, Li and Cheng in 2000. Fossils of the species were found in the village of Chuanjie, Lufeng County, Yunnan Province, (in Chuanjie Formation deposits) and are named after the location where the fossils were discovered. Известно несколько скелетов этого огромного для своего времени ящера (длина - 25 м). Недавно открытое захоронение множества зауропод в Китае неподалеку от места первоначальной находки может также содержать и эту разновидность. Имел довольно длинную шею. Обитал в батонском веке (средняя юра - 180 - 170 млн. л. н.) на территории провинции Юннань современного Китая.
Отбивает нападение аллозавров
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| 11:11 am [industrialterro] [Link] |
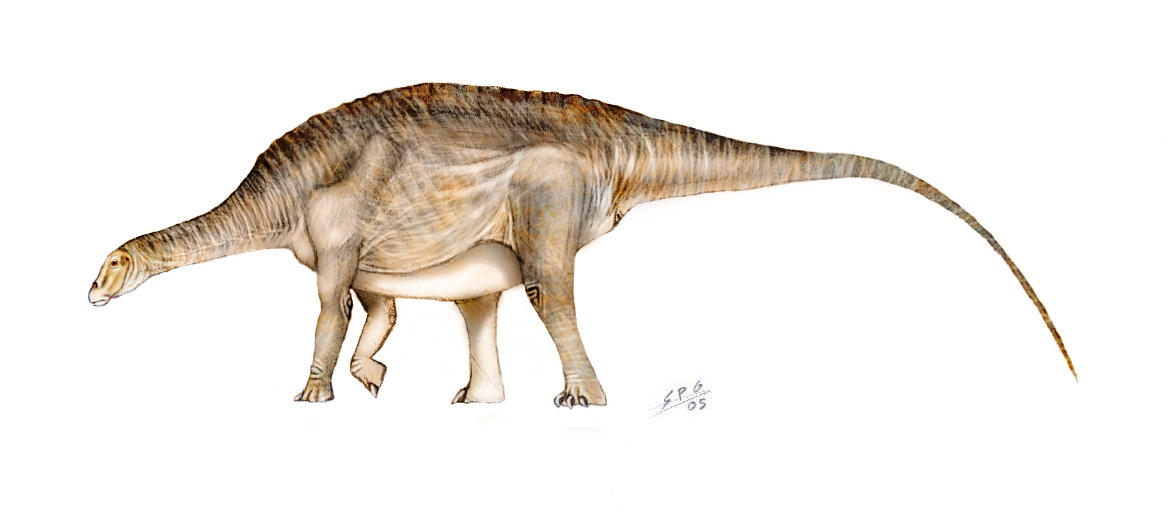
Cetiosaurus Цетиозавры (др.-греч. κῆτος — «кит» и σαῦρος — «ящерица», буквально «китоящерица») — род ящеротазовых динозавров инфраотряда зауропод, обитавший в среднеюрский период (181—169 млн лет назад). Останки известны из отложений Европы и Африки. Цетиозавр достигал 18 м в длину. Его шея была примерно такой же длины как и туловище. Хвост был значительно длиннее и включал в себя около 40 позвонков. Спинные позвонки были утяжелёнными, в отличе от полых позвонков других зауропод, таких как брахиозавр. Предплечье было более длинным, чем у других зауропод. Длина бедра цетиозавра — около 1,8 м. Cetiosaurus является первой находкой среди зауроподов, также является самым известным зауроподом из Англии. Ископаемые останки были найдены в Англии и Марокко. Окаменелости найденные в Англии состоят из позвонков, ребра и кости предплечья. Были обнаружены на острове Уайт, и назван английским биологом и палеонтологом сэром Ричардом Оуэном в 1841 году, за год до этого он ввел в обращение термин динозавры. The closest relatives of Cetiosaurus appear to be Barapasaurus and the South American Patagosaurus. Together they comprise the Cetiosauridae, which was previously a large ill-defined family of primitive sauropods. It must also be noted that Chebsaurus is often referred to the Cetiosauridae, and that Barapasaurus has been assigned to the Vulcanodontidae, due to its having cervical vertebrae that resemble other cetiosaurids, but a narrow sacrum that is a defining factor for vulcanodontids. It shared its time period with, and was possibly prey to, Megalosaurus and Eustreptospondylus. The environment in which Cetiosaurus lived was floodplain and open woodland. Названне, означающее китовый ящер, было дано этому динозавру в 1841 г., и это является одним из множества примером того, как останки динозавров путали с костями совершенно других животных. Первоначально считалось, что цетиозавр был какой-то гигантской морской рептилией, и лишь в 1869 г. цетиозавра признали динозавром. Цетиозавры были массивный животными весом до 27 тонн. У них были относительно короткие шея и хвост, но конечности впечатляли своими размерами: бедренные кости достигали почти 2 м в длину. Передние и задние конечности были примерно одинаковой длинны, а это означало, что цетиозавры держали спину горизонтально. Этим они отличались от многих поздних зауропод, у которых задние и передние конечности были разной длины. Палеонтологи не обнаружили ни одного фрагмента черепов цетиозавров, поэтому ученые не знают, чем они питались, но вероятно, зубы у них походили на грабли, что позволяло срывать листья с низкорослых деревьев и кустарников. Давая название этому ящеру, палеонтологи принимали цетиозавра за огромную, плававшую в море, рептилию. В дальнейшем он был отнесен к зауроподам. В отличие от многих других зауроподов, имевших полые кости, позвонки цетиозавра по строению относились к губчатым костям. Это утяжеляло его длинную шею и снижало ее гибкость, из-за чего он редко поднимал голову, но его окружало достаточное количество папоротников и низкорослых лиственных деревьев, чтобы он мог прокормиться. Плоские, ложкообразные зубы этого динозавра срезали листья, а камешки в желудке перетирали растительную массу для ее дальнейшего переваривания. Цетиозавр был самым первым среди найденных зауроподов. C. medius (1842) - это первый описанный зауропод, давший наименование целой группе, отличавшейся примитивным строением (сплошные, без полостей позвонки, в области таза срослись только 4 позвонка вместо 5 и даже 6 у продвинутых зауропод, короткие, но крепкие и массивные кости ног, толстые, широкие ступни, менее длинные шея и хвост). В настоящее время изучено 2 частичных скелета 15-18 м длины без черепа (известен лишь неполный зуб), а также изолированные позвоночники и элементы конечностей от множества животных по всей территории Англии. C. mogrebiensis, найденный в Марокко, отличается от C. oxoniensis относительно длинными передними лапами и тем, что все 3 найденные кости значительно стройнее, чем у английского вида, кости предплечья относительно короче, череп больше похож на черепа китайских представителей группы, минимум 13 спинных позвонков, лопатка с более расширенным концом. Это вполне может свидетельствовать о том, что этот вид можно выделить в отдельный род. Один из скелетов этого вида теперь принадлежит к отдельному роду Atlasaurus, близкому к Brachiosaurus. Известно несколько частичных скелетов этого вида, некоторые из них сохранились с черепами. Род основан на C. oxoniensis - умеренно длинная шея и относительно короткий хвост, плечевая кость равна голени, но не короче, как у большинства зауропод. Несколько отличается от ранее описанного C. medius. Цетиозавриды (Cetiosauridae) — семейство примитивных ящеротазовых динозавров из группы зауропод, живших в юрском и меловом периодах, на территории нынешней Европы и Северной Африки. Название семейству впервые предложено Лайдеккером в 1888 году. В настоящее время цетиозавры относятся к кладе Eusauropoda. Так же в некоторых классификациях, Cetiosauridae вместе с другими семействами - Vulcanodontidae и Omeisauridae не включены в группу зауропод, поскольку они считаются парафилетическими, или даже (в случае Camarasauridae) полифилетическими.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, цетиозавриды, ящеротазовые |
| 10:47 am [industrialterro] [Link] |
Cetiosauriscus Cetiosauriscus (meaning "whale-lizard-like" i.e. "Cetiosaurus-like") is a genus of sauropod dinosaur. It was perhaps a diplodocid, a close relative of Diplodocus, and lived in the Callovian (Middle to Late Jurassic Period) of England (about 162 million years ago (mya). Cetiosauriscus was a quadrupedal, herbivorous, saurischian dinosaur. Cetiosauriscus was first named by German palaeontologist Friedrich von Huene, in 1927, as the generic name of the species Cetiosauriscus leedsi. In 1887 John Whittaker Hulke had named the species Ornithopsis leedsii, based on specimen BMNH R.1984-1988, a set of bones from the Leeds collection. After a suggestion by Harry Govier Seeley, in 1905 this species was renamed to Cetiosaurus leedsi by Arthur Smith Woodward who referred a second specimen from the same collection to the species: BMNH R.3078. Both specimina were assigned to Cetiosauriscus leedsi by von Huene. In 1929 he also renamed Cetiosaurus greppini, a species found in Switzerland, Cetiosauriscus greppini. In 1980 Alan Jack Charig concluded that BMNH R.3078 could not be referred to BMNH R.1984-1988, due to a lack of comparable bones, and created a new species for the former: Cetiosauriscus stewarti. The specific name honours Sir Ronald Stewart, the chairman of the London Brick Company which owned the clay pit the fossils had been found in. Furthermore he considered both C. leedsi and C. greppini nomina dubia. In 1990 John Stanton McIntosh renamed two more species of Cetiosaurus into Cetiosauriscus longus and Cetiosauriscus glymptonensis respectively. Both were again considered nomina dubia by Charig, who in 1993 petitioned the ICZN to make Cetiosauriscus stewarti the type species instead of the original Cetiosauriscus leedsi. This was done in 1995, making BMNH R.3078, already the holotype of the species C. stewarti, the genoholotype of the genus Cetiosauriscus. BMNH R.3078 was found at Peterborough, Cambridgeshire, in strata of the Upper Oxford Clay and consists of a series of vertebrae from the rear half of the skeleton. Other remains, including a front leg, have been referred to C. stewarti. It has been estimated that this creature stood six metres high and was fifteen metres in length, weighing about nine tonnes. It may have been preyed on by carnivores such as Megalosaurus and Eustreptospondylus. Von Huene originally classified Cetiosauriscus as a member of the Cardiodontinae within the Cetiosauridae. In 1978 McIntosh concluded it belonged tot the Diplodocidae because of double chevrons under its caudal vertebrae, the short front legs and a tabular bump on the lower end of its first metatarsal. This would make Cetiosauriscus the oldest known diplodocid. Other researchers have suggested it could have belonged to the Mamenchisauridae instead, which group also features double chevrons.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, диплодоциды, завроподоморфы, завроподы, ящеротазовые |
| August 3rd, 2012 | |
| 03:20 pm [industrialterro] [Link] |
Camarasaurus Camarasaurus ( Camarasaurus is among of the most common and frequently well-preserved sauropod dinosaurs. The maximum size of the most common species, C. lentus, was about 15 meters (50 ft) in length. The largest species, C. supremus, reached a maximum length of 23 meters (75 ft) and maximum estimated weight of 47 tonnes (51.8 tons). The arched skull of Camarasaurus was remarkably square and the blunt snout had many fenestrae, though it was sturdy and is frequently recovered in good condition by paleontologists. The 19 centimeter long (7.5 in) teeth were shaped like chisels (spatulate) and arranged evenly along the jaw. The strength of the teeth indicates that Camarasaurus probably ate coarser plant material than the slender-toothed diplodocids. Each front limb bore five toes, with the inner toe having a large sharpened claw. Like most sauropods, the front limbs were shorter than the hind legs, but the high position of the shoulders meant there was little slope in the back. Serving the purpose of weight-saving, as seen in other sauropods, many of the vertebrae were hollowed out or pneumatic, riddled with passages and cavities for an intricate system of air sacs connected to the lungs. This feature was relatively unknown at the time Camarasaurus was discovered, and was the inspiration for its name, meaning "chambered lizard". The neck and counterbalancing tail were shorter than usual for a sauropod of this size. Camarasaurus, again like certain other sauropods, had an enlargement of the spinal cord near the hips. Palaeontologists originally believed this to be a second brain, perhaps necessary to co-ordinate such a huge creature. However, while it would have been an area of large nervous, probably reflex (automatic) activity, it was not a brain, and such enlargements are actually found to some degree in all vertebrate animals. Camarasaurus lewisi and C. grandis both had more robust radii than fellow sauropod Venenosaurus. There is a fossil record of two adults and a 12.2 meter (40 ft) long juvenile that died together in the Late Jurassic Period, approximately 150 million years ago (in north east Wyoming, USA, excavated by the Division of Vertebrate Paleontology of the University of Kansas Natural History Museum and Biodiversity Center, during the 1997 and 1998 'field seasons'). It is assumed that their bodies were washed to their final resting place, in alluvial mud, by a river in spate. This suggests that Camarasaurus traveled in herds or, at least, 'family' groups. Also, recovered camarasaur eggs have been found in lines, rather than in neatly arranged nests as with some other dinosaurs, which appears to suggest that, like most sauropods, Camarasaurus did not tend its young. Previously, some scientists had suggested that Camarasaurus and other sauropods may have swallowed stones (gastroliths) to help grind the food in the stomach and then regurgitated or passed them when they became too smooth. However, more recent analysis of the evidence for stomach stones has suggested that this was not the case. Furthermore, the strong, robust teeth of Camarasaurus in particular were more developed than those of most sauropods, indicating that it may have processed food in its mouth to some degree before swallowing The type species of Camarasaurus is Cope's original species, C. supremus (meaning "the biggest chambered lizard"), named in 1877. Other species since discovered include C. grandis ("grand chambered lizard") in 1877, C. lentus in 1889, and C. lewisi (originally described as a the new genus Cathetosaurus) in 1988. C. supremus, as its name suggests, is the largest known species of Camarasaurus and one of the most massive sauropods known from the late Jurassic Morrison Formation. Besides its huge size, it was nearly indistinguishable from C. lentus. C. supremus was not typical of the genus as a whole, and is only known from the latest, uppermost parts of the formation. C. grandis and C. lentus were both smaller and present in the earlier stages of the Morrison. Stratigraphic evidence suggests that the chronological sequence of the Camarasaurus species aligned with their physical differences and describe an evolutionary progression within the Morrison Formation. C. grandis is the oldest known species, present in the lowest rock layers of the Morrison. C. lentus appeared later, and co-existed with C. grandis for several million years, possibly due to different ecological niches suggested by differences in the spinal anatomy of the two species. At a later stage, C. grandis disappears from the rock record, leaving only C. lentus, which then also disappeared at the same time C. supremus appears in the uppermost layers. This immediate succession of species, as well as the very close similarity between the two, suggests that C. supremus may have evolved directly from C. lentus, representing simply a larger, later-surviving population of animals. Первое упоминание об этом ящере датировано 1877 годом, когда О. В. Лукас нашел несколько изолированных позвонков в штате Колорадо, а Эдвард Дринкер Коп дал им название. Первое описание целого скелета сделано в 1925 году Чарльзом В. Гилмором. Скелет принадлежал молодой особи и очень часто именно его и изображают на иллюстрациях. Сам камаразавр вероятно достигал длины 18 метров (60 футов). Он был растительноядным животным. Сложенная верхнеюрскими породами формация Моррисон расположена на восточных склонах Скалистых гор, где простирается с севера на юг по территории штатов Монтана и Нью-Мексико, США. На залитых водой и поросших лесом долинах и дельтах рек в поздней юре происходило накопление глин и илов этой формации, там и обитали все виды зауропод — диплодоки, апатозавры, брахиозавры и многие другие. Но ни один вид не был столь многочисленным, как камаразавр. Важнейшая отличительная особенность камаразавра — строение его черепа. Он был большим, немного походим на коробку и насчитывал много окон — больших отверстий между костями. Зубы ложковидной формы располагались по всей длине челюстей, а не только в передней их части, как у представителей подсемейства диплодоцид. Передние ноги были короче задних, как у большинства зауропод, но лопатки оставались высокими, так что спина его вряд ли наклонялась вперед. Позвонки не имели длинных отростков, поэтому маловероятно, что камаразавр поднимался на задних ногах. Шея и хвост его были короче, чем у большинства зауропод. Наличие сильных зубов говорит о том, что камаразавр питался более твердой пищей, чем диплодоциды. Скорее всего, он заглатывал камни, с помощью которых в желудке перетиралась пища. Найденные в формации Моррисон кучи отполированных камней — возможное свидетельство того, что животное, похожее на камаразавра периодически освобождалось от камней из желудка, как это делают современные куры, когда желудочные камни у них истираются (прим. - на самом деле, точное назначение камней-гастролитов для камаразавра неизвестно).
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |
| August 1st, 2012 | |
| 01:11 pm [industrialterro] [Link] |
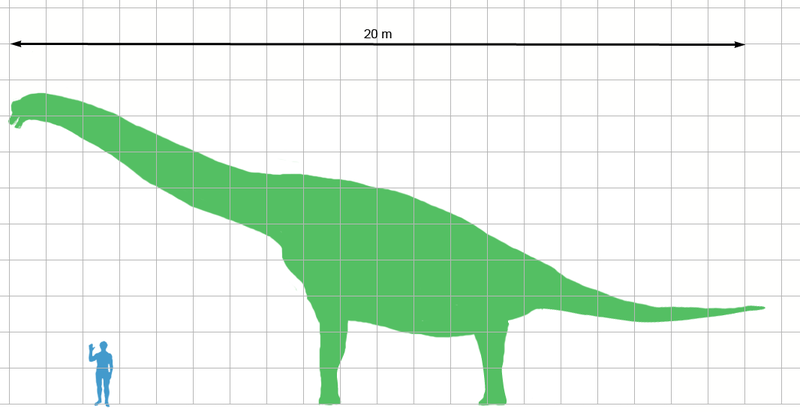
Brachiosaurus Брахиоза́вр (Brachiosaurus; буквально «плечистый ящер») — вымерший род растительноядных динозавров из группы зауроподов-макронарий, живших в конце юрского периода (около 161,2 — 145,5 млн лет назад) на территории нынешней Северной Америки и Африки. До открытия сейсмозавра, считался самым высоким динозавром. Маленькая голова на конце восьмиметровой шеи находилась на высоте 13 метров. Ноздри брахиозавра находились на своеобразном полукруглом костном гребне выше глаз. Вероятно, ноздри соединялись с воздушными мешками, как у некоторых утконосых динозавров. Также это может объяснятся тем, что у ящера, возможно, был короткий хобот. Передние ноги брахиозавра существенно длиннее задних, по внешнему виду ящер напоминал гигантского жирафа. Однако его шея была направлена не вверх, а вперед под углом примерно 45 градусов. Зубы мощные, ложковидные. Вероятно, ящеры питались листвой деревьев. Брахиозавры жили на территории Северной Америки и Африки (Танзания) в конце юрской эпохи, но сходные с ними ящеры (например, завропосейдон) дожили до середины мелового периода в Северной Америке и достигли даже более крупных размеров, чем брахиозавры. Американский брахиозавр (B. altitorax Riggs, 1903) — один из самых крупных зауропод, длиной до 27 метров и весом около 50 тонн. В Берлинском музее имени Гумбольдта выставлен скелет 14-метрового Brachiosaurus brancai (найденый германской Восточно-Африканской палеонтологической экспедицией на горе Тендагуру в Танзании), который теперь относится к отдельному роду жираффатитан. Brachiosaurus nougaredi был выделен Lapparent в 1960 году определен как новый вид брахиозавра, описаный по части крестца и передней конечности, найденых в местности Wargla в Алжире. Позже было установлено, что данные окаменелости относятся к альбскому веку, и примерно на 40 миллионов лет моложе Brachiosaurus altithorax и Giraffatitan. В настоящее время таксон помечен как nomen dubium, вероятно, из-за низкого качества ископаемого материала. В 2003 году Герхардом Майером (Gerhard Maier) в Indiana Univeristy Press был описан новый род Abdallahsaurus, который в настоящее время помечен как nomen nudum, а его останки относя к Giraffatitan. ( Далее ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14):
( Далее ) Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, брахиозавриды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| June 2nd, 2012 | |
| 06:18 pm [industrialterro] [Link] |
Brachytrachelopan Брахитрахелопан (Brachytrachelopan, от греч. "короткошеий Пан") — род динозавров, живший 150—145 млн лет назад (титонский ярус позднего юрского периода) на территории нынешней Аргентины. Достигал длины 10 метров, высоты 3 метров и веса 5—7 т. Впервые описали Оливер Раухут, Ремес, Фехнер, Кладера и Пуэрта в 2005 году. Характерной чертой брахитрахелопана является крайне короткая шея, состоящая всего из 12 позвонков. Она больше напоминает шею стегозавра, чем диплодоцида. Как и у других членов группы дикреозавров, у него были высокие шипы на стпине и бёдрах, предположительно поддерживавшие какое-то образование, похожее на парус. Шипы к передней части изгибаются вперёд. Это доказывает то, что животное держало голову книзу и питалось с земли. Череп и хвост единственного известного скелета отсутствуют. Останки B. mesai обнаружил аргентинский овчар Дэниель Меза, искавший потерявшуюся овцу. Родовое название выбрано в честь этого события (Пан — бог пастухов), видовое — по фамилии пастуха. В результате филогенетических анализов, основываясь на синапоморфии, Brachytrachelopan отнесли к семейству Dicraeosauridae надсемейства Diplodocoidea. This taxon's very short neck (approximately 40% shorter than other dicraeosaurids and the shortest of any known sauropod) is evidence that this lineage specialized to fill an ecological niche not exploited by other members of this infraorder. Small for a sauropod, Brachytrachelopan measured less than 10 metres (33 ft) in length. Rauhut et al. (2005, 670) note that the high degree of fusion present between the preserved neural arches and their respective centra, as well as fusion between the sacral centra, sacral neural arches, and sacral neural spines is evidence that the holotype does not represent a juvenile animal. Hence, the small body size is not a relic of ontogeny. Rauhut et al. (2005, 671-672) note that the tendency towards shorter-necks seen in dicraeosaurids, and most evident in Brachytrachelopan, runs counter to the lengthening of the neck seen most sauropod lineages (brachiosaurids, titanosaurs, diplodocids, etc.) and indicates that this group of sauropods was "progressively adapting for low browsing and might have been specialized on specific food sources, as has been suggested for Amargasaurus and Dicraeosaurus." Moreover, the morphology of the cervical neural arches in Brachytrachelopan would have significantly restricted dorsal flexion of neck and most likely indicates that this sauropod was specialized to a diet of plants "growing at heights of between about 1 and 2 m." Rauhut et al. (2005, 672) also suggest that diet may have been a limiting factor in body size among dicraeosaurids, and that this may have placed them in the same ecological niche as "large low-browsing iguantodontian ornithopods." It is notable that such large iguanodontians are absent from the Late Jurassic Gondwanan sediments that have produced all known fossils of dicraeosaurids, while they are abundant in similar ecosystems of the same age in North America, where dicraeosaurids are absent. This may indicate that large iguanodontians and dicraeosaurids (especially Brachytrachelopan) were ecological analogs, resulting from parallel evolution in two distantly related dinosaurian lineages. Dicraeosauridae is a family of sauropod dinosaurs known from the Late Jurassic and Early Cretaceous of Africa and South America. Currently only three genera are recognised; the Jurassic South American genus Brachytrachelopan, the Jurassic African Dicraeosaurus and the South American Early Cretaceous Amargasaurus, with its distinctive neck spines. All three animals are small by sauropod standards, with relatively short necks. McIntosh (1990) includes a number of genera within the subfamily Dicraeasaurinae of the family Diplodocidae, but many of these are now recognised as belonging to distinct families (Nemegtosauridae and Rebbachisauridae). The group is described cladistically as a stem-based taxon defined as "The most inclusive clade containing Dicraeosaurus hansemanni Janensch 1914 but not Diplodocus longus Marsh 1878" or as "all diplodocoids more closely related to Dicraeosaurus hansemanni than to Diplodocus longus (Sereno 1998 p.63, Upchurch et al. 2004 p.304, Wilson 2005 p.35).
Репродукции (1, 2, 3, 4, 5, 6, 7):
( Далее ) Размеры тела в сравнении с человеком и другими дикреозаврами:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, дикреозавры, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| 06:00 pm [industrialterro] [Link] |
Bellusaurus Bellusaurus (meaning "Beautiful lizard", from Vulgar Latin bellus 'beautiful' (masculine form) and Ancient Greek sauros 'lizard') was a small short-necked sauropod dinosaur from the Middle Jurassic which measured about 4.8 metres (16 ft) long. Its fossils were found in Shishugou Formation rocks in the northeastern Junggar Basin in China. The type (and only known) species is Bellusaurus sui, formally described by Chao in 1987. Seventeen individuals were found in a single quarry, suggesting that a herd had been killed in a flash flood. Some features suggest they may have all been juveniles. Mamenchisauridae (возможное русское название — Маменхизавриды) — вымершее семейства зауроподовых динозавров, известных с ранней юры и живших на территории нынешней Восточной Азии. Таксон впервые был определен китайскими палеонтологами C.C. Young и H.C. Chao в 1972 году, на основе типового рода — Mamenchisaurus. Так же в семейство Mamenchisauridae входят Chuanjiesaurus, Eomamenchisaurus, и Tonganosaurus. Останки Mamenchisaurus были найдены в геологической формации Shangshaximiao, около 161.2—155.7 млн. лет назад, Chuanjiesaurus около 175.6—161.2 млн. лет назад, в то время как Eomamenchisaurus был найден в формации Чжаньхэ (Zhanghe), около 175.6—161.2 млн. лет назад. Tonganosaurus жил еще раньше, в ранней юре.
Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, маменчизавриды, ящеротазовые |
| 05:07 pm [industrialterro] [Link] |
Barosaurus Барозавр (Barosaurus) — большой травоядный динозавр из семейства диплоцид, живший около 150 млн лет назад в Юрский период. Останки обнаружены в Африке, в Танзании, а также в США. Это было большое животное, весом в 30 тонн и длиной в 20 метров. В высоту достигал 12 метров — совсем, как брахиозавр. Был похож на своих сородичей, особенно на диплодока, но отличался более крупными размерами и массивным телом. Хвост у него однако был короче. И всё же у барозавра была очень длинная шея. Барозавр был растительноядным животным. Большую часть своего времени он посвящал кормёжке. Кормом барозавру служили листья с верхушек деревьев или низкорослые кустарники. Такому огромному динозавру, как барозавр, наверняка требовалось колоссальное питание. В день он мог съедать по 500 или даже по 1000 кг растительной пищи. Жили барозавры, как и все крупные травоядные динозавры, стадами. Вероятно, барозавры, как и многие другие динозавры, хорошо заботились о своём потомстве. Раньше считалось, что крупные завроподы просто оставляли яйца на произвол судьбы. Но скоро стало известно, что это совсем не так. Барозавры защищались мощным хвостом, а в случае необходимости могли пользоваться шеей или передними конечностями. От других диплодоцидов барозавры отличались более коротким и мощным хвостом и значительно более длинной и гибкой шеей. Ученые определили, что сердце этого ящера, чтобы качать кровь в мозг, должно было весить примерно полторы тонны. Впервые останки барозавра были найдены в XIX столетии. Этой находке наука обязана палеонтологу-любителю госпоже Е. Эллерман, занимавшейся раскопками в штате Южная Дакота. Виды барозавров: B. lentus - киммеридгиан, титониан (поздняя юра, 154-144 млн лет до н.э.) и аптиан, албиан (ранний мел,120-100 млн лат до н.э.), Штаты Нью-Мексико, Южная Дакота, Юта (США, Сев Америка) и Танзания (Африка). Длина - 23-27 м, вес - 15-25 т. 5 частичных скелетов без черепов, изолированные элементы конечностей. Основной вид - B.lentus - недостаёт черепа, передней части шеи, предплечья. Американская форма B.lentus является наибольшей из трех разновидностей Barosaurus; и идентична Diplodocus за исключением длинной шеи. Близок к роду Diplodocus (особенно строением конечностей), но отличается очень удлинёнными шейными позвонками (шея на треть длиннее, чем у Diplodocus, - 9 м, состоит из 16 позвонков длиной до 1м). B. africanus - поздний киммеридгиан (поздняя юра, 152-150 млн лет до н.э.), Танзания (Африка). Длина - 20 м, вес - 8,3 т, высота в плечах - 3 м, в бедрах – 4 м (плечевая кость - 0,97 м, локтевая кость – 0,74 м, лопатка – 1,34 м, бедро – 1,34 м), Более 3 частичных скелетов, несколько элементов черепа, много изолированных костей постчерепа от 41 экземпляра. Странно, но большой Brachiosaurus brancai, столь обычный в течение раннего времени tendaguru, полностью отсутствует в более поздней фауне. Вместо этого мы находим большую разновидность Barosaurus, и менее обильный материал нового животного - Tornieria. По численности Barosaurus africanus доминирует над поздней фауной tendaguru, Brachiosaurus brancai же доминировал над более ранней фауной, будучи более обычным, чем Dicraeosaurus и Tornieria вместе взятые. Но в индивидуальном количестве Barosaurus весит в два раза меньше, чем Brachiosaurus, и эти два вида вели различный образ жизни. Brachiosaurus питался высокорасположенными ветвями, а Barosaurus питался подлеском и низкими ветвями; чрезвычайно удлиненная, ползущая шея барозавра предназначалась для того, чтобы выбирать растительность среди ветвей и низких кустарников. B. gracilis - ранний - средний киммеридгиан (поздняя юра, 154 - 152 млн лет до н.э.). Танзания (Африка). Длина - 16 м, вес - 4,2 т. Много изолированных элементов конечностей от минимум 15 экземпляров. B. gracilus является самым ранним и наименьшим из Barosaurs - линии завропод, которые, как и маменчизавры, развивали необычайно длинные шеи. Хотя череп не известен, они вероятно имели тонкую голову, как у Diplodocus, со слабыми, подобными колышкам, зубами, которые, возможно, были пригодны только для того, чтобы раздевать листья или питаться мягкой растительностью, и менее вероятно, что они могли позволить животному употреблять в пищу хвою. Встречаясь с обеих сторон юрского прото-атлантического фарватера, Barosarus, возможно, развился от наследственной формы в течение позднего калловиана или раннего оксфордгиана. В сырой мегафауне тропического леса формации tendaguru была только одна эволюционная линия diplodocid - маленький африканский barosaurs. Но, в более сухой мегафауне Америки, на территории моррисоновской формации были найдены три отличительные линии диплодоцид: большой Barosaurs, Diplodocus и Apatosaurus. Первоначально, Barosaurs были относительно незначительным элементом мегафауны tendaguru, составляя 10 % идентифицированных экземпляров, и 4 % фактической биомассы мегафауны. Barosaurus намного более обычен в последующих отложениях, где составляет 50 % идентифицированных экземпляров и 40 % биомассы мегафауны. С возможным отсутствием brachiosaurs в этой области (возможно из-за изменения экологических факторов) Barosaurus увеличился в размере и числе, и стал доминирующим элементом фауны tendaguru. Barosaurus was an enormous animal, with some adults measuring more than 26 meters (85 feet) in length and weighing more than 20 metric tons (22 short tons). Barosaurus was differently proportioned than its close relative Diplodocus, with a longer neck and shorter tail, but was about the same length overall. It was longer than Apatosaurus, but its skeleton was less robust. Sauropod skulls are rarely preserved, and scientists have yet to discover a Barosaurus skull. Related diplodocids like Apatosaurus and Diplodocus had long, low skulls with peg-like teeth confined to the front of the jaws. Most of the distinguishing skeletal features of Barosaurus were in the vertebrae, although a complete vertebral column has never been found. Diplodocus and Apatosaurus both had 15 cervical (neck) and 10 dorsal (trunk) vertebrae, while Barosaurus had only 9 dorsals. A dorsal may have been converted into a cervical vertebra, for a total of 16 vertebrae in the neck. Barosaurus cervicals were similar to those of Diplodocus, but some were up to 50% longer. The neural spines protruding from the top of the vertebrae were neither as tall or as complex in Barosaurus as they were in Diplodocus. In contrast to its neck vertebrae, Barosaurus had shorter caudal (tail) vertebrae than Diplodocus, resulting in a shorter tail. The chevron bones lining the underside of the tail were forked and had a prominent forward spike, much like the closely related Diplodocus. The tail probably ended in a long whiplash, much like Apatosaurus, Diplodocus and other diplodocids, some of which had up to 80 tail vertebrae. The limb bones of Barosaurus were virtually indistinguishable from those of Diplodocus. Both were quadrupedal, with columnar limbs adapted to support the enormous bulk of the animals. Barosaurus had proportionately longer forelimbs than other diplodocids, although they were still shorter than most other groups of sauropods. There was a single carpal bone in the wrist, and the metacarpals were more slender than those of Diplodocus. Barosaurus feet have never been discovered, but like other sauropods, it would have been digitigrade, with all four feet each bearing five small toes. A large claw adorned the inside digit on the manus (forefoot) while smaller claws tipped the inside three digits of the pes (hindfoo).
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10):
( Далее ) Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, диплодоциды, завроподоморфы, завроподы, ящеротазовые |
| 04:23 pm [industrialterro] [Link] |
Barapasaurus Барапазавр (Barapasaurus) — род вымерших ящеротазовых динозавров из семейства Vulcanodontidae, относящегося к зауроподам, живших в юрском периоде (около 189—176 миллионов лет назад), на территории нынешней Азии. Окаменелости завропода были найдены в Индии. Впервые описан палеонтологом Jain и его коллегами в 1975 году. Представлен одним видом — B. tagorei. Этот динозавр был огромным четвероногим травоядным ящером с пильчатыми ложковидными зубами, у него была длинная шея и хвост. Большинство ученых считают его одним из древнейших представителей завпропод. Барапазавр, видимо, был весьма распространенным зауроподом ранней юры в Индии. Найдено более 300 образцов, представляющих различные части скелета примерно шести особей. Однако - как это часто случается при обнаружении остатков зауропод - так и не было найдено ни одного черепа и костей стопы, что создает определенные трудности при классификации. Хотя барапазавр был научно описан и классифицирован на основании тазобедренной кости, позже были обнаружены многие другие кости. Спинные позвонки имеют характерную глубокую щель для спинного мозга: эта особенность отличает его от других зауропод. Видимо, у него были длинные и стройные ноги. Зубы в форме ложки, как и у многих других зауропод, использовались для срывания зелени с ветвей. Узость тазобедренного пояса предполагает, что этот динозавр принадлежит к группе Cetiosauridae. а не к более примитивной группе Vulcanodontidae. Despite its early place in dinosaur history, Barapasaurus reached a length of about 18 metres (59 ft), and weighed about 48 tonnes (53 short tons). Its height to the hip was approximately 5.5 metres (18 ft). Like all sauropods, Barapasaurus was a herbivore. However, no skull has yet been recovered and so its exact diet has not been determined. A few lone teeth are known but not sufficient to make judgments on diet. Barapasaurus is included in Vulcanodontidae rather than, as was previously the case, the Cetiosauridae, another family in Sauropoda. This classification is unconfirmed since insufficient work has been done on the skeletons. The description is justified by the narrow sacrum (the hip vertebrae), which is a characteristic of vulcanodontids. However, the back vertebrae display features shared with Cetiosauridae, such as a high neural arch above the vertebral foramen. Currently the only known species is the type species B. tagorei. The first bones of Barapasaurus were recovered in India in 1960. However, it was not described until 1975 recovery of the type specimen was made, and the official description published by Jain, Kutty, Roy-Chowdhury and Chatterjee of Calcutta. Since then, five additional skeletons have been recovered from the Godavari Valley in southern India. However, none of these included the skull or feet, and little work has yet been published regarding these discoveries. The Early Jurassic sauropod dinosaurs Zizhongosaurus, Barapasaurus, Tazoudasaurus, and Vulcanodon may form a natural group of basal sauropods called the Vulcanodontidae. Basal vulcanodonts include some of the earliest known examples of sauropods. The family-level name Vulcanodontidae was erected by M.R. Cooper in 1984. In 1995 Hunt et al. published the opinion that the family is synonymous with the Barapasauridae. One of the key morphological features specific to the family is an unusually narrow sacrum.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, вулканодонтиды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| May 30th, 2012 | |
| 08:32 pm [industrialterro] [Link] |
Australodocus Australodocus, meaning "southern beam" from the Latin australis "southern" and the Greek dokos/δοκоς "beam", is a sauropod dinosaur that lived during the Late Jurassic period, around 150 million years ago in what was then German East Africa (now Tanzania). Its scientific name is derived from its originally being considered a southern (Gondwanan) relative of Diplodocus, though this classification is now considered unlikely. The species name honors Boheti bin Amrani, a native crew supervisor and chief preparator who was an important contributor to the German expeditions that first excavated the Tanzanian sites. The remains of Australodocus bohetii were recovered from the Tendaguru beds of Tanzania, which have been a fertile ground for many Jurassic dinosaurs, including several genera of large sauropods, such as Giraffatitan (also known as Brachiosaurus brancai), Janenschia, Tendaguria, and Tornieria. Australodocus itself is based on two neck vertebrae, which are less elongate than those of other diplodocids and differ in other anatomical details. These vertebrae were originally part of a series of four vertebrae collected in the 1909 expedition led by Werner Janensch; unfortunately, like some other fossils collected by German expeditions to Africa, the other bones were destroyed in World War II. The 2007 description of the surviving bones increases the known diversity of sauropods and diplodocids in Tendaguru. Australodocus was originally described as a diplodocid, due to the fact that it had double (bifurcate') neural spines on some of its vertebra, a characteristic normally associated with diplodocoid sauropods. However, several later studies by John Whitlock and colleagues found that Australodocus is actually a member of the sauropod clade Titanosauriformes, possibly closely related to Brachiosaurus. The presence of a higher number of macronarian sauropods in the Tendaguru environment compared to numerous diplodocoids in the Morrison Formation may be due to previously known differences in environment, with the Tendaguru being dominated by conifer forest, and the Morrison being dominated by open plains of low-browse flora. Титанозавроформы (Titanosauriformes) — группа четвероногих растительноядных динозавров отряда ящеротазовых, обитавших с юрского по меловой периоды (около 200 — 85 миллионов лет назад) на всех материках. Описанный в 2010 году динозавр Fukuititan расширяет биологическое разнообразие титанозавроформов на территории Японии и дает ученым доказательства о том, что титанозавроформы были более широко представленны на данной территории, чем до этого считалось ранее.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| 08:24 pm [industrialterro] [Link] |
Archaeodontosaurus Archaeodontosaurus ("ancient-toothed lizard") is a genus of sauropod dinosaur from the Middle Jurassic. Its fossils were found in Madagascar. The type species, Archaeodontosaurus descouensi, was described in September 2005. The specific name honours the collector, Didier Descouens. It is a probable sauropod, with prosauropod-like teeth.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| 07:39 pm [industrialterro] [Link] |
Apatosaurus Апатоза́вр ( Apatosaurus, «обманчивый ящер»), более широко известный как бронтозавр — вымерший род ящеротазовых динозавров, живших в позднеюрском периоде (около 157—146 миллионов лет назад) на территории нынешней Северной Америки. Один из крупнейших представителей зауропод. Brontosaurus excelsus был описан Гофониилом Маршем в 1879 г. по хорошо сохранившимся остаткам; впоследствии выяснилось, что он должен составлять один род с описанным тем же Маршем в 1877 году Apatosaurus ajax и по правилам ICZN родовое название Apatosaurus должно употребляться для обоих видов, как имеющее приоритет. Тем не менее, название «бронтозавр» широко укоренилось в научно-популярной литературе. Бронтозавр (апатозавр) — один из крупнейших динозавров: его длина могла достигать 20—23 метров, а масса, по разным данным, до 24—32 тонн, однако некоторые ученые указывают на то, что их масса не превышала 18 тонн. Длинные хвост и шея, массивные ноги, на конце шеи — относительно маленькая голова, мозг в которой весил всего 400 граммов. Апатозавр (более известный как бронтозавр) был одним из самых крупных из когда-либо существовавших животных. Его длина около 21-27 метров, а вес 30-35 тонн. Его голова имела небольшие размеры: менее чем 60 см в длину. Апатозавр имел продолговатый череп и маленький мозг. Травоядное имело длинную шею (с 15 позвонками) и длиный кнутообразный хвост (около 15 метров длиной). Его задние конечности были длиннее чем передние. Апатозавр мог удерживать свою шею почти горизонтально. С помощью длинной шеи он мог "скашивать" растения или проникать вглубь леса между близкорастущими растениями. Американский палеонтолог Отниел Марш описал дал имя апатозавру в 1877 году. А в 1879 году он описал и дал имя бронтозавру. Как оказалось, два динозавра были, на самом деле, двумя видами одного рода. Было признано более ранне имя - апатозавр. Черепа растительноядных завропод сохраняются довольно редко, поскольку они состоят из облегченных тонких костей и являются порой почти ажурными. Поэтому ученые часто не знают, как выглядят черепа многих завропод, хотя остальные части скелета уже обнаружены. В одном из карьеров скелетные останки апатозавра были обнаружены вместе со скелетом и черепом камаразавра. В те времена ученые еще не знали. кому какой череп принадлежал и предположили что найденный череп мог принадлежать апатозавру, поскольку к его скелету он лежал чуть ближе, чем к скелету камаразавра. Многие годы в палеонтологических музеях были выставлены скелеты апатозавра с черепом, принадлежавшим камаразавру. В результате апатозавр оставался попросту без головы, до тех пор пока не был найден его скелет с сохранившимся черепом и истина была, наконец, восстановлена. У апатозавра на шейных и спинных позвонках имелись небольшие по размеру двуглавые выросты, игравшие важную роль в строении скелета животного . Они поддерживали массивные канатоподобные сухожилия, которые в свою очередь удерживали шею и хвост. На позвонках примыкающих к тазу таких выростов не было, но сами позвонки были очень длинными. К ним крепились мощные мыщцы, связанные с конечностями животного и позволяющие апатозавру приподниматься на задних лапах. При этом хвост играл роль своеобрзной третьей ноги, помогая животному сохранять равновесие. В результате апатозавр мог дотягиваться до самых верхних нежных и сочных веточек и листьев.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10):
( Далее ) Размеры тела в сравнении с человеком (показаны разные виды апатозавров):
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, диплодоциды, завроподоморфы, завроподы, ящеротазовые |




































































































/Brachytrachelopan01.jpg)








































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.001_-_London.JPG){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
-PaleozoologicalMuseumOfChina-May23-08.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}