[Recent Entries][Archive][Friends][User Info]
Below are the 15 most recent journal entries recorded in the "Сообщество, посвящённое ра" journal:| February 11th, 2016 | |
|---|---|
| 11:06 pm [industrialterro] [Link] |
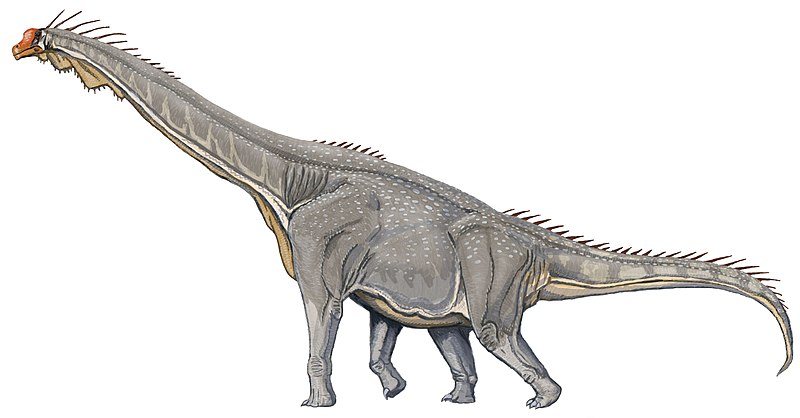
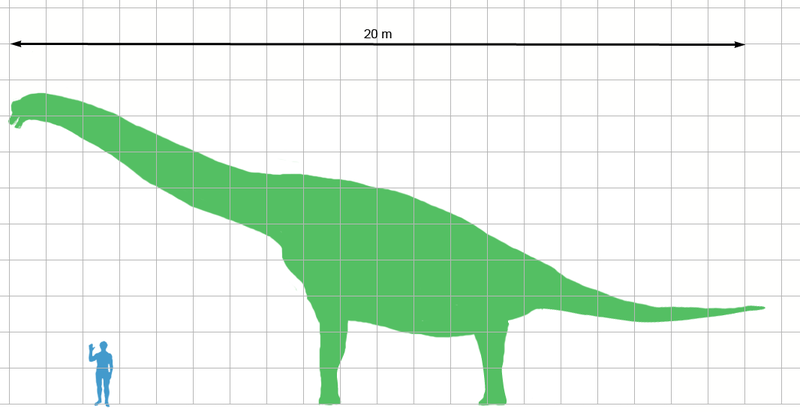
Ampelosaurus Ampelosaurus (/ˌæmpᵻloʊˈsɔːrəs/ am-pi-lo-sawr-əs; meaning "vine lizard") is a titanosaurian sauropod dinosaur hailing from the Late Cretaceous Period of what is now Europe. Its type species is A. atacis, named by Le Loeuff in 1995. A possible unnamed species has givenAmpelosaurus an age reaching to the latest Cretaceous, from about 70 to 66 million years ago. Like most sauropods, it would have had a long neck and tail but it also carried armor in the form of osteoderms. Over 500 bones have been assigned to Ampelosaurus and all but the braincase (assigned to A. sp.) has been assigned to A. atacis. They are assigned to the same species because all the differences in the limb proportions have been linked to individual variation. A. atacis is known from a few, well-preserved teeth and some cranial material. A right scapula was discovered associated with a coracoid. The blade of the scapula, contrary to most titanosaurs, is triangular. The blade narrows at one end instead of showing an expansion like most other genera. A. atacis is also known from a pubis about 75 centimetres (30 in) long and an ilium. Aside from that, it is known from a partial forelimb, and many femora. Titanosaurians were a flourishing group of sauropod dinosaurs during Cretaceous times. The Spanish locality from the latest Cretaceous of “Lo Hueco” yielded a relatively well preserved, titanosaurian braincase, which shares a number of unique features with A. atacis from France. However, it appeared to differ from A. atacis in some traits also. The specimen has been provisionally identified as Ampelosaurus sp.. A. sp. is most likely a mature titanosaurian since the bones of the braincase have largely fused together. Ampelosaurus lived alongside many other animals. Over 8500 specimens have been found alongside it, including gastropods, bivalves, crocodiles, other sauropods, plants and invertebrates in the Villalba de la Sierra, Gres de Saint-Chinian, Marnes Rouges Inférieures and Gres de Labarre formations. Recent attention has made Ampelosaurus one of the most well-known dinosaurs known from France. Like most sauropods, Ampelosaurus would have had a long neck and tail, but it also carried armor in the form of osteoderms 25 to 28 centimetres (9.8 to 11.0 in) long. The four osteoderms found have three different morphologies, they are plate, bulb, and spine-shaped. This dinosaur would have stretched up to about 15 metres (49 ft) from snout to tail. Ampelosaurus atacis is known from many unattached bones and teeth. It is one of the best-known dinosaurs from France. It is considered to be a dwarf sauropod by Coria et al. (2005) because it has a smaller size in comparison with its ancestors. Ampelosaurus is one of the most completely known titanosaurian sauropod from Europe. Since 1989, over 500 bones discovered at the Upper Cretaceous Bellevue locality of southern France have been assigned to A. atacis. An abundant amount of well-preserved elements have been uncovered at Bellevue. In the original description, only a tooth and some vertebrae were examined. In 2005, a complete description was made by Le Loueff of all known material, allowing Ampelosaurus to be much better understood. All the material has been assigned to the species, and even though there are some differences in limb proportions, it has all been linked to individual variation. A. atacis is known from a few, well-preserved teeth and some cranial material, a tooth of which was described in its original description. The teeth differentiate Ampelosaurus from Magyarosaurus and Lirainosaurus, both of which are also from Europe. The teeth are different from the two later genera as Ampelosaurus has teeth that are roughly cylindrical in the top with thin expansions at the bottom. They are 21 millimetres (0.83 in) high and about 6 millimetres (0.24 in) wide. Those expansions give its teeth a slight constriction between the root and the crown. The teeth are also quite different from the peg-like teeth of titanosaurids. The teeth, have a general morphology similar to titanosauriansAntarctosaurus and a braincase from Dongargaon. Another skull was described from southern France, but showed some differences, meaning there could have been at least two different titanosaurians in France during the Late Cretaceous. The scapula was found attached to a coracoid. In the longer direction, the bone is 72 centimetres (28 in) long. The blade of the scapula, contrary to titanosaurs, is triangular, larger closer to the base. The blade narrows distally instead of showing an expansion. Differing fromMagyarosaurus and Saltasaurus, the scapula does not have the dorsal crest at the base of the scapular blade. The shape of the coracoid is quadrangular, and the coracoid has a thickened cranial margin. Ampelosaurus is also known from a pubis, about 75 centimetres (30 in) long. It has a strong distal expansion, that is similar to a titanosaur from Brazil, and a large foramen. Contrary to the South American titanosaurs, the ilium does not have an expansion on the side. The ischium, known from a shaft, is unexpanded distally and very well developed. It is also very flat. An incomplete radius is also known. It lacks the proximal and distal ends, and possesses a very prominent ridge along it. The largest and best-preserved specimens are both ulnas. The right ulna has a total length of 395 millimetres (15.6 in). The right ulna is small and slender, is missing an olecranon, and has a well-marked ridge. The left ulna is larger, with a length of 725 millimetres (28.5 in). The ulna has a deep radial fossa. About 27 femora are known from Bellevue, making them the most common bone, most of them more-or-less complete. They are very flat craniocaudally but otherwise, lack any unusual features. In a study of its bone physiology, it was found that the maximum known femur length was 1,100 millimetres (43 in), which is much larger than previously known (802 millimetres (31.6 in)). That means, that the ancestorsAmpelosaurus were slightly smaller than itself. 18 humeri are also known, but none of them approach the size of the largest femur. The humeri do not have a free medullary cavity. The humeri are robust, about 63 centimetres (25 in) long, with very expanded distal and proximal ends. Three different types of osteoderms have been recovered at Bellevue. Two with large spines have been uncovered. At the base of each spine, which is about 12 centimetres (4.7 in) tall, are two large foramina. The internal faces are all concavo-convex and have a ridge opposite the spine. In side view, they have two sections, a low half that progressively thickens, and a spine. ( Read More ) Размеры тела в сравнении с человеком: Ископаемые останки и реплики (1, 2, 3, 4, 5, 6, 7, 8): ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| March 24th, 2015 | |
| 04:37 pm [industrialterro] [Link] |
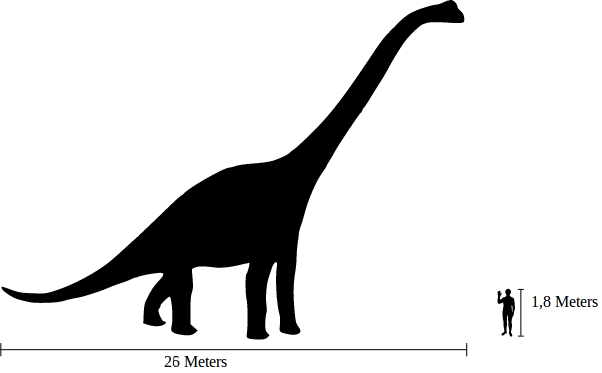
Alamosaurus Аламозавр (Alamosaurus) — род ящеротазовых динозавров инфраотряда завропод. Включает один вид — Alamosaurus sanjuanensis. Эти ящеры жили в верхнем мелу (около 70—65 млн лет назад) на территории Северной Америки. Они достигали длины 21 м и веса 30 тонн. У аламозавра, как и у других завропод, была длинная шея и не менее длинный хвост, заканчивающийся кнутоподобным сужением. Вопреки популярному утверждению, этот динозавр не был назван в честь миссии Аламо в Техасе или произошедшей там в 1836 году битвы. Голотип этого вида был обнаружен в Нью-Мексико, и только после этого другие экземпляры были найдены и в Техасе. Родовое название дано по имени геологической свиты Ojo Alamo, в породах которой нашли первый экземпляр, а видовое — по имени округа Сан-Хуан штата Нью-Мексико, где была сделана эта находка. Аламозавра уверенно относят к группе титанозавров, в пределах которой он считается продвинутым родом. Но его родственные связи в пределах этой группы остаются неясными. Некоторые авторы включают его вместе с родом Opisthocoelicaudia в подсемейство Opisthocoelicaudinae семейства Saltasauridae. Согласно другим исследованиям, аламозавр — сестринская группа рода Pellegrinisaurus, и в состав Saltasauridae они не входят. Некоторые учёные отмечают сходство аламозавра с Neuquensaurus и Trigonosaurus. Alamosaurus (meaning "Ojo Alamo lizard") is a genus of titanosaurian sauropod dinosaurs, containing a single species, Alamosaurus sanjuanensis, from the late Cretaceous Period of what is now southern North America. It was a large quadrupedal herbivore. Isolated vertebrae and limb bones indicate that it reached sizes comparable to Argentinosaurus and Puertasaurus, which would make it the largest dinosaur known from North America. Its fossils have been recovered from a variety of geological formations spanning the Maastrichtian stage (late Edmontonian-Lancian land vertebrate ages) of the Late Cretaceous. Specimens of a juvenile Alamosaurus sanjuanensis have been recovered from only a few meters below the Cretaceous-Paleogene boundary in Texas, making it among the last surviving non-avian dinosaur species. The vertebrae from the middle part of its tail had elongated centra. Alamosaurus had vertebral lateral fossae that resembled shallow depressions. Fossae that similarly resemble shallow depressions are known from Saltasaurus, Malawisaurus, Aeolosaurus, and Gondwanatitan. Venenosaurus also had depression-like fossae, but its "depressions" penetrated deeper into the vertebrae, were divided into two chambers, and extend farther into the vertebral columns. Alamosaurus had more robust radii than Venenosaurus. Long thought to have been unarmored, recent discoveries indicate that Alamosaurus was armored like other lithostrotians, including Saltasaurus. Alamosaurus remains have been discovered throughout the southwestern United States. The holotype was discovered in the Naashoibito Member of the Ojo Alamo Formation (or Kirtland Formation under a different definition) of New Mexico which was deposited during the Maastrichtian stage of the Late Cretaceous Period. Bones have also been recovered from other Maastrichtian formations, like the North Horn Formation of Utah and the Black Peaks, El Picacho and Javelina Formations of Texas. Gilmore originally described a scapula (shoulder bone) and ischium (pelvic bone) in 1922. In 1946, he found a more complete specimen in Utah, consisting of a complete tail, a right forelimb complete except for the tips of the toes, and both ischia. Since then, many other bits and pieces from Texas, New Mexico, and Utah have been referred to Alamosaurus, often without much description. The most completely known specimen is a recently discovered juvenile skeleton from Texas, which allowed educated estimates of length and mass. One specimen of Alamosaurus, "USNM 15660", was first uncovered in 1937. Although originally found by Gilmore, it was first recognized by Michael Brett-Surman. It has been catalogued under the same number as a more complete and well-known specimen of Alamosaurus, which was found in very close proximity based on bone impressions. In 2015 it was realized that the specimen, which had been previously described, also included osteoderms, the first confirmation they existed on Alamosaurus. No skull material is known, except for a few slender teeth. Contrary to popular assertions, this dinosaur is not named after the Alamo in San Antonio, Texas, or the battle that was fought there. The holotype, or original specimen, was discovered in New Mexico and, at the time of its naming, Alamosaurus had not yet been found in Texas. Instead, the name Alamosaurus comes from Ojo Alamo, the geologic formation in which it was found and which was, in turn, named after the nearby Ojo Alamo trading post (since this time there has been some debate as to whether to reclassify the Alamosaurus-bearing rocks as belonging to the Kirtland Formation or whether they should remain in the Ojo Alamo Formation). The term alamo itself is a Spanish word meaning "poplar" and is used for the local subspecies of cottonwood tree. The term saurus is derived from saura (σαυρα), Greek for "lizard" and is the most common suffix used in dinosaur names. There is one species (A. sanjuanensis), which is named after San Juan County, New Mexico, where the first remains were found. Both genus and species were named by Smithsonian paleontologist Charles W. Gilmore in 1922. Alamosaurus fossils are most notably found in the Naashoibito member of the Ojo Alamo Formation (dated to between about 69-68 million years old) and in the Javelina Formation, though the exact age range of the later has been difficult to determine. A juvenile specimen of Alamosaurus has been reported to come from the Black Peaks Formation, which overlies the Javelina in Big Bend, Texas, and which straddles the Cretaceous-Paleogene boundary. The Alamosaurus specimen was reported to come from a few meters below the boundary, dated to 66 Ma ago, though the position of the boundary in this region is uncertain. Only one geological site in the Javelina Formation has thus far yielded the correct rock types for radiometric dating. The outcrop, situated in the middle strata of the formation about 90 meters below the K-Pg boundary and within the local range of Alamosaurus fossils, was dated to 69.0 plus or minus 0.9 million years old in 2010. Using this date, in correlation with a measured age from the underlying Aguja Formation and the likely location of the K-Pg boundary in the overlying Black Peaks Formation, the Alamosaurus fauna seems to have lasted from about 70-66 million years ago, with the earliest records of Alamosaurus near the base of the Javelina formation, and the latest just below the K-Pg boundary in the Black Peaks Formation. Skeletal elements of Alamosaurus are among the most common Late Cretaceous dinosaur fossils found in the United States Southwest and are now used to define the fauna of that time and place. In the south of Late Cretaceous North America, the transition from the Edmontonian to the Lancian is even more dramatic than it was in the north. Thomas M. Lehman describes it as "the abrupt reemergence of a fauna with a superficially 'Jurassic' aspect." These faunas are dominated by Alamosaurus and feature abundant Quetzalcoatlus in Texas. The Alamosaurus-Quetzalcoatlus association probably represent semi-arid inland plains. The appearance of Alamosaurus may have represented an immigration event from South America. Some taxa may have co-occurred on both continents, including Kritosaurus and Avisaurus. Alamosaurus appears and achieves dominance in its environment very abruptly, which might support the idea that it originated following an immigration event. Other scientists speculated that Alamosaurus was an immigrant from Asia. However, critics of the immigration hypothesis note that inhabitants of an upland environment like Alamosaurus are more likely to be endemic than coastal species, and tend to have less of an ability to cross bodies of water. Further, Early Cretaceous titanosaurs are already known, so North American potential ancestors for Alamosaurus already existed. Other contemporaneous dinosaurs from that part of the world include tyrannosaurs, smaller theropods, the hadrosaurs Edmontosaurus sp. and Kritosaurus sp., the ankylosaur Glyptodontopelta, and the ceratopsids Torosaurus utahensis and Ojoceratops fowleri.
Размеры тела в сравнении с человеком:
Ископаемые останки и реплики (1, 2, 3, 4):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| March 1st, 2015 | |
| 06:57 pm [industrialterro] [Link] |
Agustinia Агустиния (Agustinia ligabuei) — вид ящеротазовых динозавров монотипного рода Agustinia из группы зауроподоморфов, живших в раннем меловом периоде (около 116—100 миллионов лет назад), на территории нынешней Южной Америки. Окаменелости были найдены в провинции Неукен (формация Lohan Cura) в Аргентине. Впервые описан палеонтологом Бонапарте в 1999 году. The name Agustinia honors the discoverer of the specimen, Agustin Martinelli. This dinosaur was originally named in a 1998 abstract written by famous Argentine paleontologist Jose Bonaparte. The original generic name was "Augustia", which, as it turned out, was already preoccupied by a beetle (see also: Megapnosaurus, Protognathosaurus). Bonaparte changed the name to Agustinia in a full paper published in 1999. There is one named species (A. ligabuei), which is named in honor of Dr. Giancarlo Ligabue, a philanthropist who provided financial support to the expedition which recovered the remains. Only fragmentary remains are known. These include fragments of vertebrae from the back, hips, and tail regions of the spinal column. Parts of the lower hind limb were also recovered: a fibula, tibia, and 5 metatarsals. A femur (thigh bone) was found at the site but was too fragmented to collect. Agustinia ligabuei was well known for its distinctive supposed armor plates, initially interpreted as a series of wide, vertical spikes and plates down the center of its back, somewhat like the unrelated Stegosaurus. However, further study of the poorly preserved fossil material showed that these "plates" were in fact more likely to be fragments of ribs and hip bones. Aside from the supposed armor, very little has been described of the anatomy of Agustinia. A fibula (lower leg bone) has been recovered that is about 3 feet (895 mm) long. When compared to the same bone in related dinosaurs, this indicates that Agustinia may have been about 50 feet (15 meters) long. However, the remains are incomplete and do not appear to have many distinctive characteristics which can be used to separate it from other sauropods. With the supposedly distinctive armor discredited, some authors have considered Agustinia a nomen dubium, based on remains not adequate to compare with related dinosaurs. Because of its supposedly unusual armor, Agustinia was originally assigned to its own family, Agustiniidae (Bonaparte, 1999). This family name has not come into wide acceptance. Agustinia is difficult to classify because of its fragmentary nature, and because it exhibits features of both diplodocoid and titanosaurian sauropods. What few distinctive features do exist in the only known specimen suggests it is a member of the clade Somphospondyli, but it is not possible to determine its relationships with any more accuracy beyond that. Somphospondyli — клада динозавров-зауроподов в составе более широкой группы Titanosauriformes. В 1998 году Пол Серено определил данную группу как состоящую из всех Titanosauriformes и более тесно связанную с Saltasaurus, чем с брахиозавром. В 2005 году Серено дал более точное определение: группа, состоящая из Saltasaurus loricatus и всех видов, более тесно связанных с Saltasaurus, чем с Brachiosaurus brancai. Название означает «губчатый позвонок» ссылаясь на синапоморфию группы. Somphospondyli по определению являются сестринской группой Brachiosauridae. Базальным членом группы вероятно является Erketu, найденный в 2006 году. Таксон по некоторым определениям является синонимом клады Titanosauria, хотя последняя не отвечает требованиям именования биологических таксонов согласно Международному кодексу филогенетических номенклатур. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, сомфоспондилы, титанозавроформы, ящеротазовые |
| February 18th, 2015 | |
| 04:02 pm [industrialterro] [Link] |
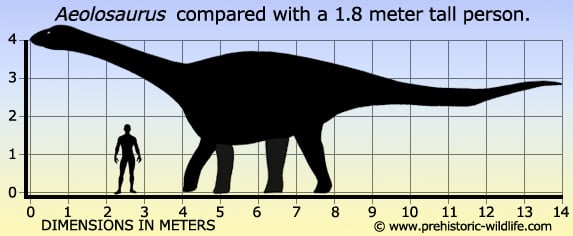
Aeolosaurus Эолозавр (Aeolosaurus) — род травоядных динозавров из группы титанозавров инфраотряда зауроподов, живший в меловом периоде около 70—68 миллионов лет назад, на территории нынешней Южной Америки. Окаменелости четвероногого зауропода были найдены в Аргентине и Бразилии. Описаны Powell в 1987 году. Всего известно 3 вида эолозавров: Aeolosaurus colhuehuapensis. Aeolosaurus maximus — вид, описанный в 2011 году. Находка была сделана в формации Adamantina на территории современной Бразилии. Aeolosaurus rionegrinus. Aeolosaurus (/ˌiː.əlɵˈsɔrəs/; "Aeolus' lizard") is a genus of titanosaurian sauropod dinosaur from the Late Cretaceous Period of what is now South America. Like most sauropods, it would have been a quadrupedal herbivore with a long neck and tail. The remains of this dinosaur are incomplete, so size can only be estimated but Aeolosaurus was probably at least 45 feet (14 m) in length. This dinosaur is named after the Greek mythological figure Aeolus, Keeper of the Winds in Homer's Odyssey, because of the frequent winds that blow across Patagonia, where the remains were found. The generic name also includes the Greek sauros ('lizard'), the traditional suffix used in dinosaur names. The specific name (A. rionegrinus), refers to its location, in the Rio Negro Province of Argentina. Both genus and species were named and described by Argentine paleontologist Jaime Powell in 1987. The holotype of Aeolosaurus rionegrinus consists of a series of seven tail vertebrae, as well as parts of both forelimbs and the right hindlimb. It was discovered in the Angostura Colorada Formation in Argentina, which dates from the Campanian stage of the Late Cretaceous, about 83 to 74 million years ago. Relationships among the many titanosaurian sauropods are hazy at best, but Aeolosaurus has been tentatively linked to a few other genera, based on features of the tail vertebrae, including Rinconsaurus and Adamantisaurus. Gondwanatitan and Aeolosaurus both exhibit neural spines on the tail vertebrae that point forwards, a feature not seen in any other known titanosaurians. The vertebrae from the middle part of its tail had elongated centra. Aeolosaurus had vertebral lateral fossae that resembled shallow depressions. Fossae that similarly resemble shallow depressions are known from Saltasaurus, Alamosaurus, Malawisaurus, and Gondwanatitan. Venenosaurus also had depression-like fossae, but its "depressions" penetrated deeper into the vertebrae, were divided into two chambers, and extend farther into the vertebral columns. Its middle tail vertebrae's neural spines angled anteriorly when the vertebrae are aligned. These vertebrae resemble those of Cedarosaurus, Venenosaurus, and Gondwanatitan. Another specimen, described in 1993, consists of five tail vertebrae, and some bones from the forelimb and pelvis. Since there are two right ulna (forearm) bones, the specimen must consist of at least two individuals. These bones were also associated with two osteoderms, or bony armor plates, providing evidence that this dinosaur was armored. This specimen is from the Allen Formation of Rio Negro, dating back about 70 to 68 million years ago to the middle Maastrichtian stage. While this specimen bears features that characterize the genus Aeolosaurus, it is from a younger time period and shows enough differences that the authors recognized it as a possible second species. Another partial skeleton, including four more tail vertebrae and material from both limbs on the left side of the body, was described in 1997. This was recovered from the Los Alamitos Formation of Rio Negro, which falls between the other two dates. This specimen was also referred to the genus Aeolosaurus, but not to the species A. rionegrinus, and may represent a third species. However, since the genus Aeolosaurus is not well known, the authors chose not to formally name either of these possible new species. For now, they are both simply known as "Aeolosaurus sp." Future discoveries may give scientists more information on variation within the genus, and show that all of the above specimens belong to A. rionegrinus, or that they merit being formally named. Another series of 15 tail vertebrae was assigned to Aeolosaurus in the original description, but it was later determined that the series does not belong to this genus, as it lacks several features found in the other specimens of Aeolosaurus. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| February 17th, 2015 | |
| 09:54 pm [industrialterro] [Link] |
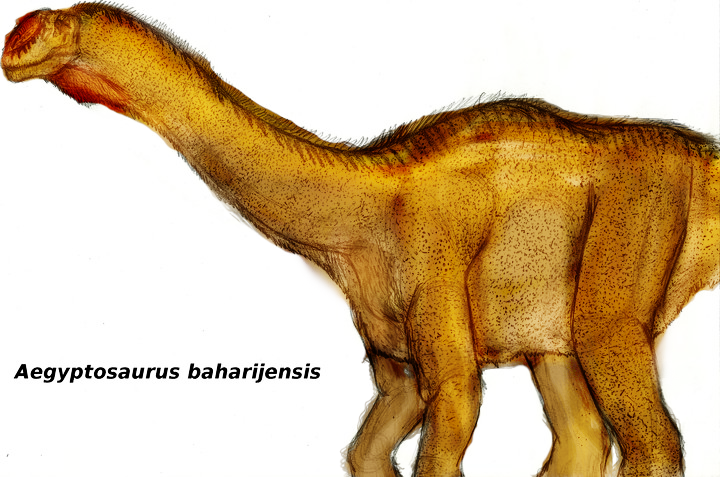
Aegyptosaurus Египтозавр (Aegyptosaurus; букв. «ящер из Египта») — род травоядных динозавров из группы титанозавров, инфраотряда зауроподов, живший в конце мелового периода (альб и сеноман) около 95 миллионов лет назад, на территории нынешней Африки. Окаменелости четвероногого зауропода были найдены в Египте, Нигере и в различных местах пустыни Сахара. Вид описан Штормером в 1932 году. Всего известно около семи скелетов Aegyptosaurus. Соотношение костей плечевой кости к бедру - 0,78, локтевой к плечевой - 0,75, голени к бедру - 0,69. Всего известно около семи скелетов египтозавра. Египтозавры по строению тела очень похожи на гигантских динозавров - представителей титанозавридов - аргентинозавров. Вполне возможно, египтозавры являются потомками аргентонозавров. Это предположение основано также на гипотезе о существовании когда-то «моста» между континентами, по которому некоторые виды древних животных расселялись по всей суше. Aegyptosaurus /iːˌdʒɪptɵˈsɔrəs/ meaning 'Egypt’s lizard', for the country in which it was discovered (Greek sauros meaning 'lizard') is a genus of sauropod dinosaur believed to have lived in what is now Africa, around 95 million years ago, during the mid- and late-Cretaceous Period (Albian to Cenomanian stages). Like most sauropods, it had a long neck and a small skull. The animal's long tail probably acted as a counterweight to its body mass. Aegyptosaurus was a close relative of Argentinosaurus, a much larger dinosaur found in South America. Aegyptosaurus was described by German paleontologist Ernst Stromer in 1932. Its fossils have been found in Egypt, Niger and in several different locations in the Sahara Desert. All known examples were discovered before 1939. The fossils were stored together in Munich, but were obliterated when an Allied bombing raid destroyed the museum where they were kept in 1944, during World War II. It is possible that Aegyptosaurus was common prey for large predatory dinosaurs, such as Carcharodontosaurus and Spinosaurus. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| February 15th, 2015 | |
| 06:12 pm [industrialterro] [Link] |
Adamantisaurus Адамантизавр (Adamantisaurus) — род динозавров из группы титанозавров, инфраотряда завроподов, живших в позднем мелового периоде, на территории современной Южной Америки. Известен только по шести хвостовым позвонкам, но, так как этот ящер является завроподом, можно предположить, что этот динозавр был очень крупным животным с длинной шеей и хвостом. Точный внешний вид восстановить невозможно, пока не будет найден более полный материал. Как и все завроподы, это был крупный, четырехногий травоядный динозавр с длинными шеей и хвостом. До сих пор науке известны лишь 6 соединенных друг с другом передних хвостовых позвонков, а также 2 нижних гемапофиза (отростки хвостовых позвонков), которые были обнаружены в бразильском штате Сан-Паулу. Род Адамантизавр, представленный единственным видом Adamantisaurus mezzalirai, впервые был описан в 2006 году бразильскими палеонтологами Сантуччи (Santucci) и Бертини (Bertini). Родовое название «Adamantisaurus» указывает на принадлежность ископаемых останков динозавра к формации Адамантина (Adamantina). Видовое название «mezzalirai» дано в честь палеонтолога Серджио Меццалиры (Mezzalira), собравшего кости и оставившего первое упоминание о них в научной литературе. Таким образом, полное название «Adamantisaurus mezzalirai» может быть переведено на русский язык как «Адамантизавр Меццалиры». В момент обнаружения 6 хвостовых позвонков были анатомически связаны между собой. Предполагается, что это – ряд хвостовых позвонков со второго по седьмой. Позвонки сильно вогнуты с передней стороны. Они имеют несколько автапоморфий (отличительных признаков), позволяющих отделить этот род от других: так, например, направленные вверх отростки позвонков чуть отогнуты назад и имеют увеличенные с боков верхние концы; постзигапофизы и презигапофизы (механические элементы соединения позвонков) тоже имеют увеличенные поверхности сочленения, причем поверхность сочленения постзигапофизов вогнута. Эти позвонки еще в 1958 году, во время строительства железной дороги из Адамантины в Ирапуру, обнаружил палеонтолог Серджио Меццалира (Sergio Mezzalira) вместе с несколькими зубами титанозавров. Место находки расположено близ города Флорида-Паулиста на юго-западе штата Сан-Паулу и относится к геологической формации Адамантина (Adamantina formation), входящей в свиту пластов Бауру (Bauru). Точный возраст свиты Бауру до сих пор оспаривается, однако авторы первого описания адамантизавра предполагают, что кости относятся к слоям от позднего кампанского яруса до раннего маастрихта. Первоначально (в 1966, 1989 годах) Меццалира приписал одной и той же особи адамантизавра не только обнаруженные позвонки, но и бедренную кость, извлеченную строителями железной дороги из того же места, где позже нашлись и позвонки. Однако авторы первого научного описания посчитали, что эта кость принадлежала, вероятно, не этому адамантизавру, а другой особи, так как она, например, относительно мала по сравнению с позвонками и отличается от них по степени сохранности. Кроме того, в формации Адамантина часто встречаются места, в которых рядом обнаруживается сразу несколько видов, что позволяет предположить, что упомянутая бедренная кость могла принадлежать другому виду. Титанозавры (Titanosauria) — группа ящеротазовых динозавров инфраотряда зауроподов, живших в юрском и меловом периодах около 171—65,5 миллионов лет назад на всех континентах планеты, кроме Антарктиды. Являлись травоядными динозаврами, обладавшими длинными шеями и хвостами, и передвигавшиеся на четырёх ногах. В длину достигали до 35—40 метров и весили между 88 и 110 тоннами. ( Read More )
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| December 23rd, 2014 | |
| 09:20 pm [industrialterro] [Link] |
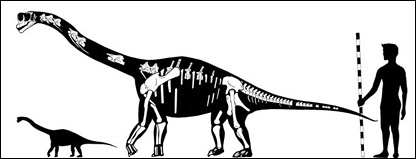
Abydosaurus Абидоза́вр (Abydosaurus; букв. «ящер Абидоса») — род ящеротазовых динозавров из семейства брахиозавриды инфраотряда зауропод, живших в раннем меловом периоде (около 105—104 млн лет назад) на территории Северной Америки (ныне штаты Юта и Колорадо, США). Представлен единственным видом — Abydosaurus mcintoshi. Первые останки динозавра, принадлежащему этому роду, были обнаружены в США на дне карьера на территории национального парка Dinosaur National Monument, на границе штатов Юта и Колорадо. В 2009 году для извлечения костей из горных пород костей динозавров на территории парка был использован динамит. Описан в 2010 году группой палеонтологов под руководством Daniel Chure. Всего палеонтологи нашли разрозненные кости скелетов четырёх молодых особей динозавров и четыре черепа, два из которых сохранились практически полностью. Длина молодых особей этого вида составляла 7-8 метров, размер взрослых представителей был, вероятно, гораздо крупнее. Название «абидозавр» является ссылкой на египетскую мифологию — Абидос в Древнем Египте был основным центром поклонения Осирису; как считалось, здесь захоронена голова бога (тогда как голотип абидозавра состоял из черепа и шеи). Данный типовой вид был назван Abydosaurus mcintoshi в честь палеонтолога Джека Макинтоша (John S. ("Jack") McIntosh). Абидозавр жил приблизительно 105 миллионов лет назад, в конце раннего мелового периода, в альбском веке. Исследование костей Abydosaurus показали его тесное родство и, вероятно, происхождение от более раннего завропода брахиозавра. В связи с этим D. J. Chure (2010) выделяет данный род в семейство Brachiosauridae. It is of interest because it is one of the few sauropods known from skull material, with the first described complete skull for a Cretaceous sauropod from the Americas. It is also notable for its narrow teeth, as earlier brachiosaurids had broader teeth. Abydosaurus is based on DINO 16488, a nearly complete skull and lower jaws with the first four neck vertebrae. Abundant skull and postcranial bones were found at the same site, including partial skulls from three additional individuals, a partial hip and associated tail vertebrae, a shoulder blade, an upper arm bone, and hand bones. These fossils were found in a sandstone bed at the base of the Mussentuchit Member of the Cedar Mountain Formation, near the old visitor center of Dinosaur National Monument. Zircons from mudstones beneath the bone-bearing sandstone indicate the age of the sandstone and the its contained bones is less than 104.46 ± 0.95 million years, in the Albian stage of the Early Cretaceous. Although Abydosaurus lived some 45 million years after Giraffatitan, - the skulls of these two genera are similar except for the narrower, sharper teeth and smaller nose of Abydosaurus. Abydosaurus can be differentiated from all other sauropods, including Giraffatitan, by subtle features of the nasal and maxillary bones, its relatively small external nares (nostrils), and some features of the teeth.
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, брахиозавриды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| October 30th, 2012 | |
| 07:50 pm [industrialterro] [Link] |
Klamelisaurus Klamelisaurus is the name given to a genus of dinosaur from the Middle Jurassic. It was a sauropod similar to Bellusaurus, of which it may actually be an adult specimen. It was found in the Gobi desert of Asia. The type species, K. gobiensis, was described by Zhao Xijin in 1993 from the Shishugou Formation. Klamelisaurus was an essentially advanced sauropod that also shows many primitive features, and may represent an intermediate form in sauropod evolution. It had massive , spatulate teeth and high neural spine s on its backbone. It may be the adult form of Bellusaurus. Euhelopodidae is a family of sauropod dinosaurs which includes the genus Euhelopus. The family name was first proposed by American paleontologist Alfred Sherwood Romer in 1956. The four genera Chiayusaurus, Omeisaurus, Tienshanosaurus, and Euhelopus were the original proposed euhelopodines (subfamily Euhelopodinae). Other Chinese genera, such as Mamenchisaurus, used to be placed within this family, but are now in Mamenchisauridae. However, the traditional Euhelopodidae has been abandoned by many taxonomists for the time being. There may be such a family, but currently Jurassic and Cretaceous Asian sauropods are poorly known and have not yet been subjected to many detailed analyses, so it is unknown if there really is a Euhelopodidae of more than Euhelopus.
Ископаемые останки (реплики) - 1, 2, 3:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, эухелоподиды, ящеротазовые |
| October 23rd, 2012 | |
| 07:30 pm [industrialterro] [Link] |
Haplocanthosaurus Haplocanthosaurus (meaning "simple spined lizard") is a genus of sauropod dinosaur. Two species, H. delfsi and H. priscus, are known from incomplete fossil skeletons. It lived during the late Jurassic period (Kimmeridgian stage), 155 to 152 million years ago. The type species is H. priscus, and the referred species H. delfsi was discovered by a young college student named Edwin Delfs in Colorado. Haplocanthosaurus specimens have been found in the very lowest layer of the Morrison Formation, along with Hesperosaurus, Eobrontosaurus, and Allosaurus jimmadensi. Haplocanthosaurus was one of the smallest sauropods of the Morrison. While some Morrison sauropods could reach lengths of over 20 meters (or over 70 feet), Haplocanthosaurus wasn't nearly as large, and reached a total length of 14.8 meters (49 feet) and an estimated weight of 12.8 metric tons. There are four known specimens of Haplocanthosaurus: one of H. delfsi, and three of H. priscus. Of these, the type of H. delfsi is the only one complete enough to mount. The mounted specimen of H. delfsi now stands in the Cleveland Museum of Natural History, albeit with a completely speculative replica skull, as the actual skull was not recovered. Present in stratigraphic zones 1, 2, and 4. Haplocanthosaurus priscus was originally named Haplocanthus priscus by John Bell Hatcher in 1903. Soon after his original description, Hatcher came to believe the name Haplocanthus had already been used for a genus of acanthodian fish (Haplacanthus, named by Louis Agassiz in 1845), and was thus preoccupied. Hatcher re-classified his sauropod later in 1903, giving it the new name Haplocanthosaurus. However, the name was not technically preoccupied at all, since there was a variation in spelling: the fish was named Haplacanthus, not Haplocanthus. While Haplocanthus technically remained the valid name for this dinosaur, Hatcher's mistake was not noticed until many years after the name Haplocanthosaurus had become fixed in scientific literature. When the mistake was finally discovered, a petition was sent to the ICZN (the body which governs scientific names in zoology), which officially discarded the name Haplocanthus and declared Haplocanthosaurus the official name (ICZN Opinion #1633). Originally described as a "cetiosaurid", José Bonaparte decided in 1999 that Haplocanthosaurus differed enough from other sauropods to warrant its own family, the Haplocanthosauridae. Phylogenetic studies have failed to clarify the exact relationships of Haplocanthosaurus with any certainty. Studies have variously found it to be more primitive than the neosauropods, a primitive macronarian (related to the ancestor of more advanced forms such as Camarasaurus and the brachiosaurids), or a very primitive diplodocoid, more closely related to Diplodocus than to titanosaurs, but more primitive than rebachisaurids.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |
| October 22nd, 2012 | |
| 08:43 pm [industrialterro] [Link] |
Giraffatitan Жираффатитан (Giraffatitan brancai) — гигантский динозавр-зауропод из семейства брахиозавров. Описан в 1914 году Яненшем под названием Brachiosaurus brancai из позднеюрских (киммеридж) отложений Тендагуру на территории нынешней Танзании. Скелет был смонтирован в Берлинском музее Естественной истории и является самым высоким смонтированным скелетом динозавра. В отдельный род выделен Дж. Ольшевским в 1991 г., но это выделение не получило широкого признания и чаще новый род рассматривается в качестве синонима рода Brachiosaurus. Выделение рода жираффатитан обусловлено отличиями в строении черепа и скелета. От брахиозавра отличается также относительно лёгким телосложением. При длине около 25 метров весил лишь 15—30 тонн. Высота могла превышать 15 метров, но по современным данным, брахиозавры не могли держать шею вертикально. Таким образом, реальная высота головы над землёй была меньше. Высота в плечах у крупных особей составляла около 7 метров. В отличие от американского брахиозавра, известен по многочисленным скелетам разного размера (вёл стадный образ жизни). Характерен для нижних и средних слоёв отложений Тендагуру. Вероятно, питался листвой деревьев. Гребень на черепе выше, чем у американского вида, мог служить для терморегуляции или вокализации (на гребне могли быть закреплены кожистые голосовые мешки). Вряд ли умел хорошо плавать (как показано на старых реконструкциях) ввиду неуклюжести жирафоподобного телосложения и узких подошв стоп и кистей. В целом известен лучше, чем американский вид, часто изображается под названием «брахиозавр». Giraffatitan was a sauropod, one of a group of four-legged, plant-eating dinosaurs with long necks and tails and relatively small brains. It had a giraffe-like build, with long forelimbs and a very long neck. The skull had a tall arch anterior to the eyes, consisting of the bony nares, a number of other openings, and "spatulate" teeth (resembling chisels). The first toe on its front foot and the first three toes on its hind feet were clawed. Traditionally, the distinctive high-crested skull has been seen as a characteristic of the genus Brachiosaurus to which Giraffatitan brancai was originally referred, but because within the traditional Brachiosaurus material it is known only from Tanzanian specimens now assigned to Giraffatitan, it is possible that Brachiosaurus altithorax did not show this feature. For many decades, Giraffatitan was claimed to be the largest dinosaur known, (ignoring the possibly larger but lost Amphicoelias) but in the later part of the 20th century several giant titanosaurians (Argentinosaurus, Puertasaurus and Futalognkosaurus) appear to have surpassed Giraffatitan in terms of sheer mass. However, Giraffatitan, along with Brachiosaurus are still the largest dinosaurs known from relatively complete material. All size estimates for Giraffatitan are based on the specimen HMN SII, a subadult individual between 21.8–22.5 metres (72–74 ft) in length. Mass estimates are more problematic and historically have varied from as little as 15 tonnes (17 short tons) to as much as 78 tonnes (86 short tons). These extreme estimates are now considered unlikely due to flawed methodologies. More recent estimates based on models reconstructed from bone volume measurements, which take into account the extensive, weight-reducing air sac systems present in sauropods, and estimated muscle mass, are in the range of 23–37 tonnes (25–41 short tons). However, HMN SII is not the largest specimen known (an assertion suported by its subadult status) but HMN XV2, represented by a tibia 13% larger than the corresponding material on HMN SII, which might have attained 26 metres (85 ft) in length. Like other sauropods, Giraffatitan had a relatively small brain, when its massive body size is taken into account, of about 300 cm³. A 2009 study calculated its brain-to-body mass ratio (a rough estimate of possible intelligence) at a low 0.62 or 0.79, depending on the size estimate used. Giraffatitan is also similar to other sauropods in having an enlargement of the spinal cord above the hips, which some older sources misleadingly referred to as a "second brain". The placement of Giraffatitan nostrils has been the source of much debate with Witmer (2001) describing in Science the hypothesized position of the fleshy nostrils in Giraffatitan in as many as five possible locations. There has also been the hypothesis of various sauropods, such as Giraffatitan, possessing a trunk. The fact that there were no narrow-snouted sauropods (Giraffatitan included) tends to discredit such a hypothesis. Stronger evidence for the absence of a trunk is found in the teeth wear of Giraffatitan, which shows the kind of wear that would result from biting and tearing off of plant matter rather than purely grinding, which would be the result of having already ripped the leaves and branches off with its trunk. If Giraffatitan was endothermic (warm-blooded), it would have taken an estimated ten years to reach full size, if it were instead poikilothermic (cold-blooded), then it would have required over 100 years to reach full size. As a warm-blooded animal, the daily energy demands of Giraffatitan would have been enormous; it would probably have needed to eat more than ~182 kg (400 lb) of food per day. If Giraffatitan was fully cold-blooded or was a passive bulk endotherm, it would have needed far less food to meet its daily energy needs. Some scientists have proposed that large dinosaurs like Giraffatitan were gigantotherms. The nostrils of Giraffatitan, like the huge corresponding nasal openings in its skull, were long thought to be located on the top of the head. In past decades, scientists theorized that the animal used its nostrils like a snorkel, spending most of its time submerged in water in order to support its great mass. The current consensus view, however, is that Giraffatitan was a fully terrestrial animal. Studies have demonstrated that water pressure would have prevented the animal from breathing effectively while submerged and that its feet were too narrow for efficient aquatic use. Furthermore, new studies by Lawrence Witmer (2001) show that, while the nasal openings in the skull were placed high above the eyes, the nostrils would still have been close to the tip of the snout (a study which also lends support to the idea that the tall "crests" of brachiosaurs supported some sort of fleshy resonating chamber).
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, брахиозавриды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| October 18th, 2012 | |
| 07:18 pm [industrialterro] [Link] |
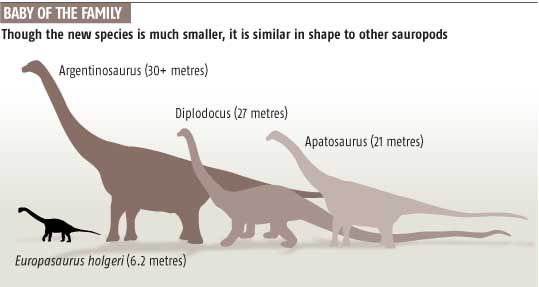
Europasaurus Europasaurus is a basal macronarian sauropod, a form of quadrupedal herbivorous dinosaur. It lived during the Late Jurassic (Kimmeridgian, between 155.7 and 150.8 million years ago) of northern Germany, and has been identified as an example of insular dwarfism resulting from the isolation of a sauropod population on an island within the Lower Saxony basin. Europasaurus holgeri exhibits the following unambiguous autapomorphic states: "nasal process of premaxillary projecting anterodorsally; medial notch is posterior dorsal margin of cervical vertebral centra; scapular acromion with a prominent posterior projection; and transverse width of astragalus twice its dorsoventral height and anteroposterior width." In comparing Europasaurus with Camarasaurus, Europasaurus differs "in the wing-shaped posterior process of the postorbital being slightly longer and wider than the anterior process, whereas it is much shorter in Camarasaurus." Other differences include Europasaurus' shorter nasal-frontal contact and a parietal bone which is rectangular in posterior view. Unlike Camarasaurus, the neural spines of the presacral vertebrae are undivided. In comparing Europasaurus with Brachiosaurus, the former genus differs from the latter by having a shorter muzzle, a quadratojugal which makes contact with the squamosal, and an anteromedially flattened humerus possessing unaligned proximal and distal epiphyses. Europasaurus was also compared with the macronarians Lusotitan and "Cetiosaurus" humerocristatus and found to be distinct from both those forms. Finally, Europasaurus can be distinguished from most known members of the Neosauropoda by its very small adult body size. Remains of both adults and juveniles have been recovered from marine carbonate beds ("bed 93"), representing more than eleven individuals ranging from 1.7 to 6.2 meters in total body length. The genus name means "lizard from Europe" (Europe + Greek sauros = lizard), and the species is named in honor of Holger Lüdtke, who discovered the first fossils of the taxa. The holotype specimen (DFMMh/FV 291; Dinosaurier-Freilichtmuseum Münchehagen/Verein zur Förderung der Niedersächsischen Paläontologie) consists of portions of a disarticulated skull, along with cervical and sacral vertebrae, derived from a single individual. This specimen and all others referred to the taxon were collected from "bed 93" at the Langenberg quarry, Oker near Goslar, Lower Saxony (Niedersachsen). A cladistic analysis of Europasaurus holgeri indicates that the species' dwarfism is a derived characteristic, and that this taxon is a more derived macronarian than Camarasaurus, and should be regarded as the sister group of the Brachiosauridae and all more derived members of the Titanosauriformes. It has been suggested that the ancestor of Europasaurus would have "dwarfed rapidly" after immigrating to a paleo-island, as even the largest of these islands in the Lower Saxony basin (a landmass <2,000 square kilometers) would not have possessed sufficient food for a population of large sauropods. Other examples of insular dwarfism among dinosaurs have been cited from the Maastrichtian (latest Cretaceous) Haţeg Island in Romania, namely the sauropod Magyarosaurus and the hadrosaur Telmatosaurus. In these two examples, paleogeography and paleoecology have been employed to explain the evolution of new and miniature versions of larger species. These fossils were determined to represent a dwarf species, and not merely juveniles of a large macronarian species, by studying the histology of the fossil bones. Likewise, by comparing the long-bone histology of a typical large-bodied sauropod, Camarasaurus, paleontologists have concluded that the diminutive size of Europasaurus resulted from a decreased growth rate. This phenomenon is a reversal of the accelerated growth responsible for gigantism in other sauropod dinosaurs. Dwarfism among macronarians seems especially remarkable as this clade includes some of the largest known dinosaurs, such as the brachiosaurs Brachiosaurus and Sauroposeidon.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11):
( Далее ) Размеры тела в сравнении с человеком (впереди неполовозрелая особь):
Размеры тела в сравнении с некоторыми другими завроподами (закрашен чёрным):
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |
| August 3rd, 2012 | |
| 03:20 pm [industrialterro] [Link] |
Camarasaurus Camarasaurus ( Camarasaurus is among of the most common and frequently well-preserved sauropod dinosaurs. The maximum size of the most common species, C. lentus, was about 15 meters (50 ft) in length. The largest species, C. supremus, reached a maximum length of 23 meters (75 ft) and maximum estimated weight of 47 tonnes (51.8 tons). The arched skull of Camarasaurus was remarkably square and the blunt snout had many fenestrae, though it was sturdy and is frequently recovered in good condition by paleontologists. The 19 centimeter long (7.5 in) teeth were shaped like chisels (spatulate) and arranged evenly along the jaw. The strength of the teeth indicates that Camarasaurus probably ate coarser plant material than the slender-toothed diplodocids. Each front limb bore five toes, with the inner toe having a large sharpened claw. Like most sauropods, the front limbs were shorter than the hind legs, but the high position of the shoulders meant there was little slope in the back. Serving the purpose of weight-saving, as seen in other sauropods, many of the vertebrae were hollowed out or pneumatic, riddled with passages and cavities for an intricate system of air sacs connected to the lungs. This feature was relatively unknown at the time Camarasaurus was discovered, and was the inspiration for its name, meaning "chambered lizard". The neck and counterbalancing tail were shorter than usual for a sauropod of this size. Camarasaurus, again like certain other sauropods, had an enlargement of the spinal cord near the hips. Palaeontologists originally believed this to be a second brain, perhaps necessary to co-ordinate such a huge creature. However, while it would have been an area of large nervous, probably reflex (automatic) activity, it was not a brain, and such enlargements are actually found to some degree in all vertebrate animals. Camarasaurus lewisi and C. grandis both had more robust radii than fellow sauropod Venenosaurus. There is a fossil record of two adults and a 12.2 meter (40 ft) long juvenile that died together in the Late Jurassic Period, approximately 150 million years ago (in north east Wyoming, USA, excavated by the Division of Vertebrate Paleontology of the University of Kansas Natural History Museum and Biodiversity Center, during the 1997 and 1998 'field seasons'). It is assumed that their bodies were washed to their final resting place, in alluvial mud, by a river in spate. This suggests that Camarasaurus traveled in herds or, at least, 'family' groups. Also, recovered camarasaur eggs have been found in lines, rather than in neatly arranged nests as with some other dinosaurs, which appears to suggest that, like most sauropods, Camarasaurus did not tend its young. Previously, some scientists had suggested that Camarasaurus and other sauropods may have swallowed stones (gastroliths) to help grind the food in the stomach and then regurgitated or passed them when they became too smooth. However, more recent analysis of the evidence for stomach stones has suggested that this was not the case. Furthermore, the strong, robust teeth of Camarasaurus in particular were more developed than those of most sauropods, indicating that it may have processed food in its mouth to some degree before swallowing The type species of Camarasaurus is Cope's original species, C. supremus (meaning "the biggest chambered lizard"), named in 1877. Other species since discovered include C. grandis ("grand chambered lizard") in 1877, C. lentus in 1889, and C. lewisi (originally described as a the new genus Cathetosaurus) in 1988. C. supremus, as its name suggests, is the largest known species of Camarasaurus and one of the most massive sauropods known from the late Jurassic Morrison Formation. Besides its huge size, it was nearly indistinguishable from C. lentus. C. supremus was not typical of the genus as a whole, and is only known from the latest, uppermost parts of the formation. C. grandis and C. lentus were both smaller and present in the earlier stages of the Morrison. Stratigraphic evidence suggests that the chronological sequence of the Camarasaurus species aligned with their physical differences and describe an evolutionary progression within the Morrison Formation. C. grandis is the oldest known species, present in the lowest rock layers of the Morrison. C. lentus appeared later, and co-existed with C. grandis for several million years, possibly due to different ecological niches suggested by differences in the spinal anatomy of the two species. At a later stage, C. grandis disappears from the rock record, leaving only C. lentus, which then also disappeared at the same time C. supremus appears in the uppermost layers. This immediate succession of species, as well as the very close similarity between the two, suggests that C. supremus may have evolved directly from C. lentus, representing simply a larger, later-surviving population of animals. Первое упоминание об этом ящере датировано 1877 годом, когда О. В. Лукас нашел несколько изолированных позвонков в штате Колорадо, а Эдвард Дринкер Коп дал им название. Первое описание целого скелета сделано в 1925 году Чарльзом В. Гилмором. Скелет принадлежал молодой особи и очень часто именно его и изображают на иллюстрациях. Сам камаразавр вероятно достигал длины 18 метров (60 футов). Он был растительноядным животным. Сложенная верхнеюрскими породами формация Моррисон расположена на восточных склонах Скалистых гор, где простирается с севера на юг по территории штатов Монтана и Нью-Мексико, США. На залитых водой и поросших лесом долинах и дельтах рек в поздней юре происходило накопление глин и илов этой формации, там и обитали все виды зауропод — диплодоки, апатозавры, брахиозавры и многие другие. Но ни один вид не был столь многочисленным, как камаразавр. Важнейшая отличительная особенность камаразавра — строение его черепа. Он был большим, немного походим на коробку и насчитывал много окон — больших отверстий между костями. Зубы ложковидной формы располагались по всей длине челюстей, а не только в передней их части, как у представителей подсемейства диплодоцид. Передние ноги были короче задних, как у большинства зауропод, но лопатки оставались высокими, так что спина его вряд ли наклонялась вперед. Позвонки не имели длинных отростков, поэтому маловероятно, что камаразавр поднимался на задних ногах. Шея и хвост его были короче, чем у большинства зауропод. Наличие сильных зубов говорит о том, что камаразавр питался более твердой пищей, чем диплодоциды. Скорее всего, он заглатывал камни, с помощью которых в желудке перетиралась пища. Найденные в формации Моррисон кучи отполированных камней — возможное свидетельство того, что животное, похожее на камаразавра периодически освобождалось от камней из желудка, как это делают современные куры, когда желудочные камни у них истираются (прим. - на самом деле, точное назначение камней-гастролитов для камаразавра неизвестно).
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |
| August 1st, 2012 | |
| 01:11 pm [industrialterro] [Link] |
Brachiosaurus Брахиоза́вр (Brachiosaurus; буквально «плечистый ящер») — вымерший род растительноядных динозавров из группы зауроподов-макронарий, живших в конце юрского периода (около 161,2 — 145,5 млн лет назад) на территории нынешней Северной Америки и Африки. До открытия сейсмозавра, считался самым высоким динозавром. Маленькая голова на конце восьмиметровой шеи находилась на высоте 13 метров. Ноздри брахиозавра находились на своеобразном полукруглом костном гребне выше глаз. Вероятно, ноздри соединялись с воздушными мешками, как у некоторых утконосых динозавров. Также это может объяснятся тем, что у ящера, возможно, был короткий хобот. Передние ноги брахиозавра существенно длиннее задних, по внешнему виду ящер напоминал гигантского жирафа. Однако его шея была направлена не вверх, а вперед под углом примерно 45 градусов. Зубы мощные, ложковидные. Вероятно, ящеры питались листвой деревьев. Брахиозавры жили на территории Северной Америки и Африки (Танзания) в конце юрской эпохи, но сходные с ними ящеры (например, завропосейдон) дожили до середины мелового периода в Северной Америке и достигли даже более крупных размеров, чем брахиозавры. Американский брахиозавр (B. altitorax Riggs, 1903) — один из самых крупных зауропод, длиной до 27 метров и весом около 50 тонн. В Берлинском музее имени Гумбольдта выставлен скелет 14-метрового Brachiosaurus brancai (найденый германской Восточно-Африканской палеонтологической экспедицией на горе Тендагуру в Танзании), который теперь относится к отдельному роду жираффатитан. Brachiosaurus nougaredi был выделен Lapparent в 1960 году определен как новый вид брахиозавра, описаный по части крестца и передней конечности, найденых в местности Wargla в Алжире. Позже было установлено, что данные окаменелости относятся к альбскому веку, и примерно на 40 миллионов лет моложе Brachiosaurus altithorax и Giraffatitan. В настоящее время таксон помечен как nomen dubium, вероятно, из-за низкого качества ископаемого материала. В 2003 году Герхардом Майером (Gerhard Maier) в Indiana Univeristy Press был описан новый род Abdallahsaurus, который в настоящее время помечен как nomen nudum, а его останки относя к Giraffatitan. ( Далее ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14):
( Далее ) Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, брахиозавриды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| May 30th, 2012 | |
| 08:32 pm [industrialterro] [Link] |
Australodocus Australodocus, meaning "southern beam" from the Latin australis "southern" and the Greek dokos/δοκоς "beam", is a sauropod dinosaur that lived during the Late Jurassic period, around 150 million years ago in what was then German East Africa (now Tanzania). Its scientific name is derived from its originally being considered a southern (Gondwanan) relative of Diplodocus, though this classification is now considered unlikely. The species name honors Boheti bin Amrani, a native crew supervisor and chief preparator who was an important contributor to the German expeditions that first excavated the Tanzanian sites. The remains of Australodocus bohetii were recovered from the Tendaguru beds of Tanzania, which have been a fertile ground for many Jurassic dinosaurs, including several genera of large sauropods, such as Giraffatitan (also known as Brachiosaurus brancai), Janenschia, Tendaguria, and Tornieria. Australodocus itself is based on two neck vertebrae, which are less elongate than those of other diplodocids and differ in other anatomical details. These vertebrae were originally part of a series of four vertebrae collected in the 1909 expedition led by Werner Janensch; unfortunately, like some other fossils collected by German expeditions to Africa, the other bones were destroyed in World War II. The 2007 description of the surviving bones increases the known diversity of sauropods and diplodocids in Tendaguru. Australodocus was originally described as a diplodocid, due to the fact that it had double (bifurcate') neural spines on some of its vertebra, a characteristic normally associated with diplodocoid sauropods. However, several later studies by John Whitlock and colleagues found that Australodocus is actually a member of the sauropod clade Titanosauriformes, possibly closely related to Brachiosaurus. The presence of a higher number of macronarian sauropods in the Tendaguru environment compared to numerous diplodocoids in the Morrison Formation may be due to previously known differences in environment, with the Tendaguru being dominated by conifer forest, and the Morrison being dominated by open plains of low-browse flora. Титанозавроформы (Titanosauriformes) — группа четвероногих растительноядных динозавров отряда ящеротазовых, обитавших с юрского по меловой периоды (около 200 — 85 миллионов лет назад) на всех материках. Описанный в 2010 году динозавр Fukuititan расширяет биологическое разнообразие титанозавроформов на территории Японии и дает ученым доказательства о том, что титанозавроформы были более широко представленны на данной территории, чем до этого считалось ранее.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| May 28th, 2012 | |
| 08:28 pm [industrialterro] [Link] |
Abrosaurus Аброзавр (Abrosaurus) — род динозавров из группы зауропод-макронарий, живших в раннем юрском периоде (около 168—161 миллионов лет назад) на территории нынешней Азии, один из многих динозавров, которрые были найдены в карьере Dashanpu провинции Сычуань, Китай. Как и большинство завроподов, Abrosaurus был четвероногим травоядным динозавром, но был довольно мал по сравнению со своими сородичами - не более чем 9 метров в длину. Голова его имела квадратную форму с закругленными боками и была увенчана небольшим костлявым гребнем, у основания которого находились ноздри. Род содержит единственный вид — А. dongpoi, и назван в честь китайского поэта одиннадцатого века Су Ши, также известного как Су Дунпо, который родился в провинции Сычуань. Макронары (Macronaria) — клада в составе группы динозавров-завроподов, известных от средней юры (батониан) до позднего мелового периодов и живших на территории нынешних Северной Америки, Южной Америки, Европы, Азии и Африки. Название буквально означает «большие ноздри», ссылаясь на большие носовые отверстия, которые находились высоко на черепе и представляли из себя резонирующих камеры. Macronaria состоит из двух основных групп: семейство Camarasauridae и клада Titanosauriformes. Titanosauriformes в свою очередь, содержит Brachiosauridae и группу Titanosauria, и является одной из крупнейших групп завроподов, которая также содержит самых больших динозавров известных науке. Зауропо́ды или завропо́ды (Sauropoda — букв. «ящероногие динозавры») — большая группа четвероногих растительноядных динозавров отряда ящеротазовых, обитавших с юрского по меловой периоды (около 200–85 миллионов лет назад) на всех материках; включала около 130 видов, до 13 семейств, около 70 родов. Наиболее известными представителями зауропод являются диплодоки, брахиозавры, апатозавры. Зауроподы представляли из себя гигантских четвероногих динозавров с небольшим, по сравнению с общим размером, черепом, массивным телом и длинной шеей, доходящей у некоторых видов до 9–11 метров. Ноздри смещены к глазницам. Зубы мелкие, шпателевидные. Вес этих гигантов колебался от 15 до 35 тонн. Одним из крупнейших представителей данной группы является сейсмозавр, достигавший длины до 36 метров. Зауроподы отличаются относительно небольшой головой, около 1/200 по сравнению с общими размерами. По сравнению с останками других частей скелета, череп зауроподов является довольно редкой находкой для палеонтологов. Известны менее одной трети черепов от общего количества родов зауроподов, и еще меньше известны находки полного черепа. Поэтому каждая находка черепа зауропода представляет для палеонтологов особый интерес. В результате открытия нового вида абидозавра в 2010 году, с отлично сохранившимся черепом, была подтверждена теория уменьшения величины зубов с ранней юры до мелового периода. Вопреки названию («ящероногие»), строением конечностей похожи не на ящериц, а на слонов (сходные адаптации к гигантизму). Более длинные передние, чем задние, ноги многих видов свидетельствуют о том, что пищу эти рептилии собирали преимущественно выше уровня туловища — кормились древесной листвой. Хотя многие другие виды могли кормиться водной и околоводной растительностью или кустарниками. Видоизменённые позвонки крестцового отдела позвоночника образовали подобие черепа, куда переместилась большая часть мозга — крестцовый мозг превышал размерами головной в 20 раз. Хвост использовался в брачных ухаживаниях: самцы били хвостами о землю, создавая оглушающий (для слуха современного человека) грохот — самки добивался самый громкий. В точке хвоста, где скорость превышала скорость звука, позвонки для прочности срослись. Удары хвостом могли также использоваться для защиты от хищников. Многие зауроподы несли на себе всевозможные шипы, гребни и прочее, что визуально увеличивало их размер и, вероятно, служило для общения с сородичами — указывало на видовую принадлежность, могло сделать их внешне крупнее и/или сообщать о достижении рептилией половой зрелости. Для защиты от хищников большая часть «украшений», если не все, была не пригодна. Зауроподы были растительноядными. Долгое время они считались полуводными животными, несмотря на полное отсутствие в скелете признаков, специфичных для водных животных. Гипотетическое использование шеи в качестве шноркеля невозможно, так как при этом грудная клетка должна была выдерживать давление воды. По ранним версиям длинная шея зауропод считалась приспособлением для кормления листьями высоких деревьев. Контраргументом такой гипотезы являются расчёты Р. Сеймура (2009): чтобы иметь возможность поднимать голову на высоту кроны дерева, артериальное давление в организме должно быть настолько высоким, что на его поддержание уходила бы приблизительно половина всей потребляемой зауроподом энергии. Учёные считают, что зауроподы (как и другой отряд ящеротазовых динозавров — тероподы) появились на территории Гондваны (например, Antetonitrus, найденный в Южной Африке) и позже расселились по всем континентам. Благодаря ископаемым находкам известно, что зауроподы 200 миллионов лет назад уже обитали на земле и развивались параллельно с прозауроподами. Одними из самых ранних и примитивных зауроподов позднетриасового периода являются — Isanosaurus, найденный в Таиланде, и Вулканодон найденный в Зимбабве, обитавшие по разные стороны океана Тетис. Это свидетельствует о том, что зауроподы имели длинную эволюционную историю в течение позднего триасового периода. Около 190 млн. лет назад зауроподы заселили территорию Индии (Barapasaurus, Kotasaurus), которая в то время была ещё частью континента Гондвана. Примерно в это же время они начали заселять другие территории, в частности территорию современной Южной Африки. Ископаемые останки говорят о том, что примерно 190—180 млн. лет назад зауроподы уже обитали в Китае (Zizhongosaurus, Kunmingosaurus), в Европе (Ohmdenosaurus) и Марокко (Tazoudasaurus). На территорию Северной Америки зауроподы мигрировали значительно позднее. Известно, что в поздний триасовый период тероподы уже населяли территорию Северной Америки, однако следов зауроподов в этот период там обнаружено не было. Единого мнения учёных по этому вопросу нет, некоторые высказывают предположение что их, скорее всего, не устраивал климат. Наибольшего разнообразия зауроподы достигли в среднеюрский период, дав множество различных видов. К концу юрского периода видовое разнообразие начинает несколько снижаться. В начале мелового периода (около 145–140 млн. лет назад) зауроподы уже населяли территорию почти всех континентов — Африки (брахиозавр), Северной Америки (апатозавр, диплодок), Европы (Xenoposeidon) и Азии (Qiaowanlong). В позднемеловом периоде происходит настоящий всплеск развития завроподов (особенно на южных континентах, где конкуренция с орнитоподами была минимальной). Основное разнообразие в этот период представляла группа титанозавров.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, ящеротазовые |





























































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}