[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 40 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| December 4th, 2011 | |
|---|---|
| 02:13 pm [industrialterro] [Link] |
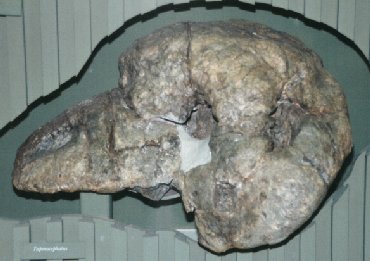
Tapinocephalus Tapinocephalus is a genus of large herbivorous dinocephalian that lived during the Middle Permian Period. These stocky, barrel-bodied animals were characterised by a massive bony skull roof and short weak snout. It is thought that, like the rest of the members of its family, the animals engaged in head-butting intraspecific behavior, possibly for territory or mates. The fossil remains (skull and postcranial elements) of Tapinocephalus are known from the Lower, Middle, and Upper part of the Tapinocephalus Assemblage Zone ( Capitanian age) of the Lower Beaufort Beds of the South African Karoo. Only the type species, T. atherstonei is now considered valid for this genus. In life, these animals were over 3 meters (10 ft) in length and weighed around 1.5 to 2 metric tonnes (1.6 to 2 short tons), making them among the largest animals of their time. Keratocephalus and Moschops are closely related contemporary forms, of similar size and habits. Тапиноцефалы (Tapinocephalia) — подотряд растительноядных дейноцефалов. Известны из средней перми Южной Африки, России и Китая. Включают два надсемейства — титанозухи и собственно тапиноцефалы. Сюда же может относиться проблематичный род стиракоцефал (Styracocephalus) из средней перми Южной Африки, которого иногда объединяют с бурнетиидами. Собственно тапиноцефалы (надсемейство Tapinocephaloidea) включает единственное семейство Tapinocephalidae. Это очень крупные животные (весом до 1 −1,5 тонн). Тело массивное, округлое, хвост очень короткий. Конечности очень массивные, передние чуть длиннее задних. Вероятно, при жизни передняя половина тела была приподнята (возможно, это указывает на питание какой-то приподнятой над почвой растительностью). Морда при обычном положении головы опущена несколько вниз (затылок резко скошен назад). Череп очень массивный, с чрезвычайно развитым пахиостозом в надглазничной и лобно-теменной области. Толщина костей черепа у некоторых видов достигала 30 см. Глазницы и височные впадины небольшие, височные впадины открыты сверху. Зубы немногочисленные, резцы обычно очень мощные. Небных зубов нет. Клыки выражены лишь у самых примитивных форм. Предполагается, что пахиостоз костей черепа был необходим для внутривидовых схваток (как у баранов или горных козлов). Тем не менее, это оспаривается отечественными учеными, которые указывают, что кости лба не были покрыты роговыми образованиями и служили для терморегуляции (их пронизывали кровеносные сосуды). Шкура тапиноцефалов, вероятно, была мягкой и лишенной чешуй, с многочисленными железами. На лицевой части черепа у некоторых видов обнаружены желобчатые углубления, возможно, вмещавшие кожные железы. Известны отпечатки следов из Южной Африки. Тапиноцефалы были растительноядными, причем для измельчения пищи они использовали мощные резцы, снабженные пяткообразными выступами. Основу их питания могли составлять мертвые стволы каламитов, которые накапливались по берегам водоемов. Самый примитивный тапиноцефал — Tapinocaninus pamelae из зоны Eodicynodon Южной Африки. Он сохранял заметные верхние клыки, но в остальном очень сходен с более поздними родичами. Найденный в Приуралье улемозавр представляет родственную примитивную форму. Недавно показано, что приуральские дейтерозавры также являются примитивными тапиноцефалами. Более прогрессивные тапиноцефалы включают примерно 12 — 13 родов преимущественно из Южной Африки. Истинное количество родов неизвестно, так как многие из них оказались синонимами, поскольку описывались на основании остатков разнополых и разновозрастных особей одного и того же вида. Tapinocephalinae — короткомордые тапиноцефалы с относительно слабыми зубами. Пахиостоз чрезвычайно развит, иногда в виде куполообразных выростов на крыше черепа. Наиболее известны мосхопс (Moschops), тапиноцефал (Tapinocephalus), кератоцефал (Keratocephalus). Самый крупный вид — Tapinocephalus atherstonei, длиной до 4 метров. Мосхопса и его ближайших родичей (Criocephalus, Avenantia, Delphinognathus) иногда выделяют в отдельное подсемейство.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 02:02 pm [industrialterro] [Link] |
Sycosaurus Sycosaurus is an extinct genus of Gorgonopsia. It was first named by Haughton in 1924, and contains four species, S. brodiei, S. kingoriensis, S. laticeps, S. vanderhorsti. Sycosaurus's skull is about medium size for a Gorgonopsid. It has a dental formula of i5, c1, pc5.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 01:47 pm [industrialterro] [Link] |
Suminia Суминия (Suminia getmanovi) — доисторический синапсид, относившийся к кладу Anomodontia, живший около 260 млн лет назад в позднем пермском периоде («зона II»). Известный экземпляр был обнаружен в г. Котельнич на реке Вятка в песчаниковых отложениях: последнее говорит о том, что скорее всего суминия была жителем речной дельты. Возможно, животное обитало и в других зонах, однако других окаменевших останков не сохранилось. Зубы обнаруженного экземпляра были сильно изношены, что говорит о том, что она питалась растительной пищей с большим содержанием частиц песка. Образцы, найденные позднее, говорят о том, что суминия вела древесный образ жизни. Единого взгляда на систематическое положение суминии нет. Наталия Рыбчински в работе, посвященной анатомии черепа и филогенетической позиции Suminia getmanovi, помещает род Suminia в инфраотряд Venyukovioidea Watson & Romer, 1956. М. Ф. Ивахненко рассматривает суминию в рамках семейства Galeopidae Broom, 1912 (инфраотряд Dromasauria, подотряд Dicynodontia). Вид Suminia getmanovi найден в местонахождении Котельнич, Котельничский р-н, Кировской обл. (верхняя пермь, татарский отдел, верхнесеверодвинский подъярус, северодвинский горизонт). Входит в состав котельничской фауны (котельничский фаунистический комплекс). Типовой экземпляр — полный скелет хорошей сохранности. В дальнейшем было найдено множество экземпляров, в том числе крупное скопление скелетов (захоронение из останков 15 животных). Ископаемые остатки нередки, представлены несколькими скелетами и черепами, а также изолированными зубами. Остатки суминий неустановленной видовой принадлежности и определенные как S. cf. getmanovi известны и из других местонахождений. Зубные кости, определенно принадлежащие роду Suminia, известны из местонахождения Потеряха-2, Нюксенский р-н, Вологодская обл. (северодвинский горизонт, верхнесеверодвинский подъярус, татарский отдел, верхняя пермь). Предчелюстная кость, отличающаяся деталями строения зубов от типового вида, найдена в местонахождении Устье Стрельны, Великоустюгский р-н, Вологодская обл. (северодвинский горизонт, верхнесеверодвинский подъярус, татарский отдел, верхняя пермь). Совершенно неожиданный взгляд на образ жизни суминии высказан в одной из недавних работ. Йорг Фребиш (Jörg Fröbisch) из Музея Филда в Чикаго (Field Museum in Chicago) и Роберт Рейц (Robert Reisz) из Университета Торонто в Канаде (University of Toronto), исследовав окаменелые скелеты суминии, обнаружили у этого аномодонта несколько адаптаций к древесному образу жизни. Рейц и Фребиш установли, что суминия была, возможно, древесным животным, и что она является на сегодняшнее время самым ранним известным древесным позвоночным. Она поднялась на деревья за 30 миллионов лет до похожих на хамелеонов дрепанозаврид, занимавших аналогичную нишу. Исследованный палеонтологами материал состоял из одного блока, содержащего остатки скелетов более десяти животных. Хотя такие захоронения могут быть трудными для изучения, они также чрезвычайно информативны, особенно с учетом возможности изучения различий между разными животными одного вида из одного места. Скелеты на плите принадлежали животным разного возраста, от полувзрослых до взрослых, а сохранность костей свидетельствует о быстром захоронении, вызванным какой-то небольшой катастрофой. Древесные животные редки в ископаемой летописи, поскольку они не живут в местах с благоприятными условиями для образования окаменелостей, поэтому ископаемые остатки суминии являются неожиданностью для палеонтологов. Прекрасная сохранность материала позволила Рейцу и Фребишу изучить анатомию суминии во всех деталях. По мнению ученых, наиболее любопытной особенностью являются кисти и стопы животных. Кисть и стопа большие по сравнению с остальной конечностью. Кисть суминии сильно отличается от кисти наземных аномодонтов и больше похожа на кисти других древесных позвоночных из разных групп и периодов времени. Большая часть длины кисти и стопы образована длинными тонкими пальцами с удлиненными предпоследними фалангами, которые по пропорциям и форме сходны с таковыми у некоторых современных животных, включая шерстокрылов, крыланов и ящериц, цепляющихся когтями при лазании по деревьям. Первые пальцы кисти и стопы могли отгибаться под углом к остальным четырем пальцам, и, возможно, использовались как противопоставленные хватательные пальцы («большие пальцы»). Это механизм, появление которого можно было бы ожидать у животного, обхватывающего конечностями ветви деревьев при перемещении в кроне и, по-видимому, кисти и пальцы суминии в этом отношении аналогичны кистям и пальцам некоторых древесных животных. Более мелкие особенности также указывают на древесный образ жизни. Хвост суминии относительно длинный, а хвостовые позвонки первой половины хвоста несут сильные отростки. Эти отростки, возможно, поддерживали мускулы, которые позволяли суминии использовать хвост для равновесия или как цепкий хватательный орган. Оказалось, что линии, которые стали вести древесный образ жизни развивали аналогичные приспособления неоднократно, и это сходство объясняется конвергентной эволюцией. Сравнение суминии с другими древесными позвоночными животными подтверждает эту гипотезу, таким образом, Рейц и Фребиш нашли доказательства того, что суминия могла перемещаться по деревьям, цепляясь и обхватывая ветви. Почему предки суминии стали жить на деревьях, однако, пока остается неясным. Суминия, жившая 260 млн. лет назад, является первым известным позвоночным животным с такой степенью специализации к древесному образу жизни. Рейц и Фребиш отмечают, что поздняя пермь и местонахождение Котельнич, где была найдена суминия, предоставляют свидетельства наиболее раннего этапа становления современных наземных экосистем с большим количеством потребителей растений, поддерживающим малое количество главных хищников. В конце пермского периода окружающая среда обеспечивала существование множества растительноядных животных (около 83% всех наземных позвоночных) и гораздо меньшего числа плотоядных (около 13% всех наземных позвоночных). Хотя древесный образ жизни помогал суминиям спасаться от хищников, авторы статьи предполагают, что именно конкуренция с другими растительноядными заставила некоторых синапсид подняться на деревья. Эти растения были ресурсом, не использовавшимся другими позвоночными, что, возможно, и предопределило эволюцию суминии от наземных предков. Таким образом, суминия нашла новый способ использовать листву верхушек деревьев и сделала первый известный шаг в нишу, к которой возвратятся позвоночные животные еще несколько раз за следующие 260 миллионов лет. В местонахождении Котельнич были найдены небольшие копролиты овально-яйцевидной формы, длиной 1-2 см, с углистым материалом внутри. В копролитах обнаружены фрагменты кутикулы, возможно, принадлежавшей птеридоспермам (семенным папоротникам), а также фрагменты древесины. Эти копролиты могли принадлежать Suminia getmanovi. Блок со скелетами суминий, изученный Фребишем и Рейцем, также содержал мелкие (менее 10 мм) копролиты, внутри которых обнаружены фрагменты листьев.
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 01:12 pm [industrialterro] [Link] |
Styracocephalus Стиракоцефал (Styracocephalus platyrhynchus) – примитивный терапсид из средней перми (средний горизонт зоны Tapinocephalus) Южной Африки. Обычно считается тапиноцефалом, но некоторыми исследователями относится к бурнетиидам. Животное крупных размеров (длиной, вероятно, до 2,5 метров). Морда очень длинная, низкая. Мощные выросты над глазами, развитые «рога» табулярных костей, направленные назад, боковые выросты на чешуйчатых костях. Зубной край верхней челюсти прямой, нижняя челюсть массивная с высоким симфизом. Развиты мощные резцы, сохраняются увеличенные клыки, заклыковые зубы многочисленные (до 11 пар). Небные зубы слабо развиты. Животное растительноядное. Описан С. Хоутоном в 1929 году. Долгое время был известен лишь по одному разрушенному черепу, но в 1997 году было опубликовано дополнительное описание еще нескольких черепов. В отличие от бурнетиид и эстемменозухов, выросты черепа образованы другими костями. Поэтому, вероятно, представляет особую ветвь дейноцефалов. С другой стороны, недавно стиракоцефала вновь сблизили с эстемменозухами и бурнетиаморфами.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| November 29th, 2011 | |
| 09:57 pm [industrialterro] [Link] |
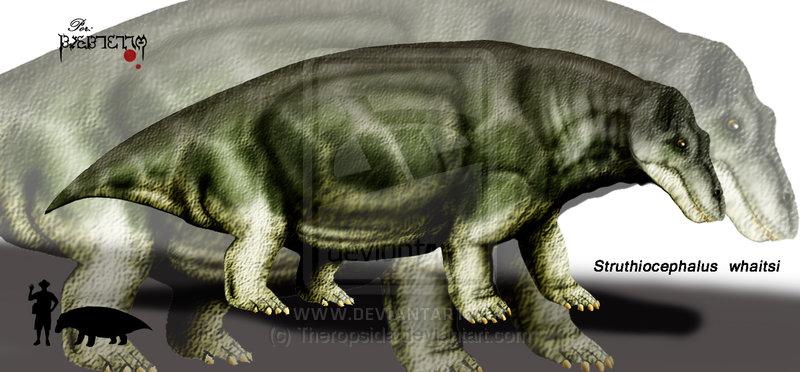
Struthiocephalus Struthiocephalus ("Ostrich Head") is an extinct genus of dinocephalian therapsid from the Permian of South Africa. Тапиноцефалы (Tapinocephalia) — подотряд растительноядных дейноцефалов. Известны из средней перми Южной Африки, России и Китая. Включают два надсемейства — титанозухи и собственно тапиноцефалы. Сюда же может относиться проблематичный род стиракоцефал (Styracocephalus) из средней перми Южной Африки, которого иногда объединяют с бурнетиидами. Струтиоцефалы (Strutiucephalinae) — длинномордые коротконогие тапиноцефалы. Пахиостоз развит значительно, переход от лицевой части к крыше черепа плавный. Возможно, полуводные животные (не исключается наличие мышечных клапанов вокруг ноздрей). 4 рода, наиболее известен струтиоцефал (Struthiocephalus), до 4 метров длиной.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:50 pm [industrialterro] [Link] |
Stenocybus Стеноцибус (Stenocybus acidentatus) – один из самых примитивных известных дейноцефалов, близкий к группе антеозавров. Обнаружен в «среднепермских» отложениях Дашанкоу, в Китае. Найденная там фауна может примерно соответствовать по возрасту Очерской фауне из Пермской области России. Длина черепа стеноцибуса около 10 – 15 см. Череп очень высокий, полукруглый (внешне напоминает черепа пеликозавров –хаптодусов). Пахиостоза нет. Мощные длинные резцы, снабженные "пяткой"; саблевидные длинные верхние клыки. Многочисленные (около 9 -10 пар) заклыковые зубы. Очень крупные глазницы. Височная впадина меньше, чем глазница. Вероятно, не относится к предкам других дейноцефалов (вместе с ним найдены остатки более продвинутых антеозавров, сходных с титанофонеусом). Может быть молодой особью крупного антеозавра синофонеуса (Sinophoneus), известного из этого же местонахождения.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:41 pm [industrialterro] [Link] |
Scylacosuchus Сцилакозух (Scylacosuchus orenburgensis) – примитивный хищный тероцефал позднепермской эпохи. Обнаружен в Оренбургской области в отложениях Ильинского субкомплекса (верхнетатарский подъярус), описан Л.А. Татариновым в 1968 г. Современник завроктона и пробурнетии. Череп длинный, высокий в передней части, 2 пары верхних «клыков» (между двумя крупными клыками – пара более мелких зубов), 6-7 пар заклыковых зубов. Вторичное небо не развито. Длина черепа 21 см. Вместе с черепом обнаружены кости почти полного скелета. Вероятно, активный хищник и падальщик. Обычно относится к семейству пристерогнатид либо сцилакозаврид (наиболее примитивные тероцефалы), но в ряде классификаций сближается с вайтсиидами.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:39 pm [industrialterro] [Link] |
Scylacosaurus Сцилакозавр (Scylacosaurus sclateri) — некрупный примитивный хищный тероцефал позднепермской эпохи. Длина черепа 18 см. Череп низкий, довольно узкий. Сохраняются заклыковые зубы (примерно 8 пар). Мощные резцы и клыки. Вторичное небо не развито. Нижняя челюсть низкая и длинная, в ней преобладает зубная кость. Известен из зоны Cistecephalus конца поздней перми Южной Африки. Вероятно, представляет реликтовую ветвь примитивных тероцефалов (относится к примитивному семейству Scylacosauridae).
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:23 pm [industrialterro] [Link] |
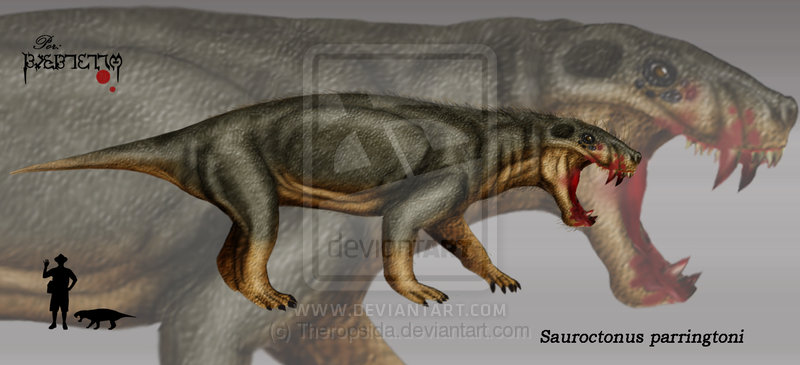
Sauroctonus Завроктон (Sauroctonus) — некрупный горгонопс из поздней перми Среднего Поволжья и Танзании. Входит в состав фауны Ильинского субкомплекса, предшествующего по времени знаменитым Северодвинским отложениям. Длина черепа у восточноевропейского Sauroctonus progressus — 225 мм (преорбитальная длина 110 мм), у южноафриканского Sauroctonus ("Aelurognathus") parringtoni — 250 мм (преорбитальная длина 135 мм). Череп европейского вида довольно узкий и низкий, у африканского вида относительно выше, с более широкой мордой. Профиль черепа несколько выгнут сверху, особенно у африканского вида, понижающийся кпереди. Височные ямы среднего размера, удлиненные, скуловые дуги узкие, глазницы небольшие. Носовые и верхнечелюстные кости сильно скульптурированы. Предтеменная кость у S. progressus очень маленькая. Отверстие теменного органа хорошо выражено. Передние части хоан расширены и образуют вместилища для крупных клыков нижней челюсти. Заклыковых зубов относительно много. Зубная формула: I 5|5, C 1|1, Pc 5|6. Имеются ряды и группы мелких тупых зубов на крыловидных, небных костях и сошнике. Небные бугры высокие, зубы на них практически редуцированы и заменены бугристыми утолщенными образованиями. Нижняя челюсть мощная, с хорошо выраженным выступом-«подбородком», венечный отросток тонкий. Африканский вид отличается более массивным челюстным симфизом. Атлант состоит из самостоятельного гипоцентра и двух несросшихся невральных дуг. В шейных позвонках присутствуют интерцентры. Конечности длинные, частично выпрямленные. Типовой вид — S. progressus. Выделен в отдельный род в 1955 году А. П. Быстровым (описан А. П. Гартман-Вейнберг в 1938 году как вид африканского рода Arctognathus). В 1940 году И.А. Ефремов относил его к роду иностранцевия. Длина черепа 225 мм, преорбитальная длина 110 мм, ширина 150 мм. Череп относительно узкий, височное окно удлиненное, орбиты некрупные, скуловые дуги узкие. Небные бугры высокие, озубленные. Африканский завроктон. В 2007 году выяснилось, что южноафриканский горгонопс Aelurognathus parringtoni, найденный в грабене Рухуху в Танзании, относится к роду Sauroctonus. Вид был описан Ф. фон Хюне в 1950 году как Scymnognathus parringtoni. В 1970 году Д. Сигонье отнесла его к роду Aelurognathus. От типового вида отличается более высокой и широкой мордой, с массивным нижнечелюстным симфизом. Длина черепа 25 см, преорбитальная длина 13,5 см, ширина затылка 14 см. Таким образом, завроктоны населяли и Северное, и Южное полушария. Завроктон — активный специализированный полуводный или наземный хищник. Анализ посткраниального скелета, выполненный Е. Гебауэр, выявил возможность передвижения горгонопсов с полувыпрямленными передними конечностями. Как и другие горгонопсы, завроктон некоторыми особенностями строения и адаптациями напоминал саблезубых кошек и, возможно, занимал сходную экологическую нишу. У обеих групп наблюдается увеличение клыков и развитие своеобразных «ножен» для клыков на нижней челюсти, сокращение числа и размера заклыковых зубов и целый ряд анатомических особенностей, позволяющих широко раскрывать пасть для укуса. Добычей завроктона могли становиться мелкие парейазавры (например, Proelginia) и дицинодонты (Tropidostoma, Idelesaurus, Dicynodon).
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:00 pm [industrialterro] [Link] |
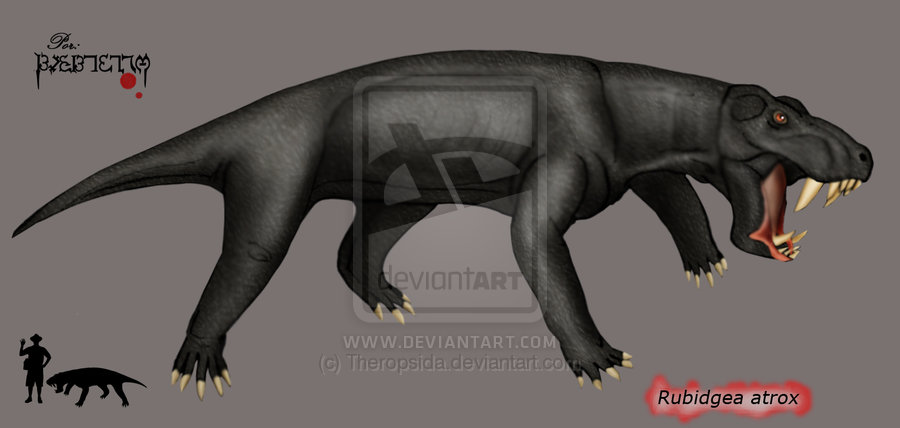
Rubidgea Рубиджея (Rubidgea) — крупный горгонопс конца позднепермской эпохи (зона Cistecephalus — Dicynodon). Один из последних крупных горгонопсов. Известны 3−5 видов, жившие только в Южной Африке. Череп относительно низкий, с большими височными отверстиями, широким затылком, передняя часть черепа высокая, скуловые дуги широкие, слегка загнуты вниз. Крыша черепа толстая, бугристая (пахиостоз). Как и у других горгонопсов, мощные резцы и огромные саблевидные клыки сочетались с практически полным отсутствием заклыковых зубов (у типового вида Rubidgea atrox — 1-2 пары, зубы исчезали с возрастом); на нижней челюсти их нет вовсе. Длина черепа доходила до 50 см и более, общая длина — до 3 м. Посткраниальный скелет адекватно не описан и плохо известен. Существующие изображения основаны преимущественно на скелете лиценопса. Статуя рубиджеи в натуральную величину есть в экспозиции музея Центра исследования ископаемых им. Китчинга в Нью-Бетесде (ЮАР). Исходя из пропорций лиценопса, длина рубиджеи могла доходить до 3 метров. Согласно ревизии Е. Гебауэр, род включает следующие виды: Rubidgea atrox — типовой вид, описан Р. Брумом в 1938 году. Длина типового черепа 47 см. Череп часто изображался в литературе, но, вероятно, был исходно сильно деформирован. Более современные реконструкции показывают большую высоту задней части нижней челюсти. Синонимы - Rubidgea platyrhina, Rubidgea majora. Происходит из терминальных пермских слоев Южной Африки. Rubidgea quinquemolaris — вид описан фон Хюне в 1950 году, как Dinogorgon quinquemolaris из поздней перми Танзании. Длина черепа 41 см. Есть пять пар заклыковых зубов на верхней челюсти, череп уже, чем у типового вида. Синоним - Dinogorgon oudebergensis. Rubidgea pricei — вид, описанный Брумом и Джорджем в 1950 году как Tigrisaurus pricei. Послеклыковых зубов нет, орбиты слабо перекрыты сверху (то есть, пахиостоз меньше развит), длина черепа 32 см. Ранее его относили также к роду Dinogorgon. К особому виду рода может принадлежать череп, описанный Р. Брумом в 1936 году как Dinogorgon rubidgei. Длина черепа могла превышать 50 см (длина ростральной части до орбит — 26 см). Есть указание на обнаружение в 1998 году в поздней перми Южной Африки гигантского рубиджеида с черепом 75 см длиной (30 дюймов), предварительно отнесенного к роду Rubidgea. В отличие от иностранцевий, рубиджеи могли быть наземными хищниками, специализированными к охоте за крупными дицинодонтами. Интересно, что в Южной Африке горгонопсы (именно рубиджея) дожили до самого конца перми, тогда как в Восточной Европе иностранцевии исчезли несколько ранее конца перми. Не исключено существование какого-то очень крупного представителя рубиджеид в поздней перми (соколковский комплекс) Вологодской области. М. Ф. Ивахненко упоминает о наличии в коллекции ПИН фрагментов черепа горгонопса, сходного с рубиджеей: длина полного черепа могла достигать 70 см. В 2003 году этот вид был назван Leogorgon klimovensis.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| November 16th, 2011 | |
| 10:42 pm [industrialterro] [Link] |
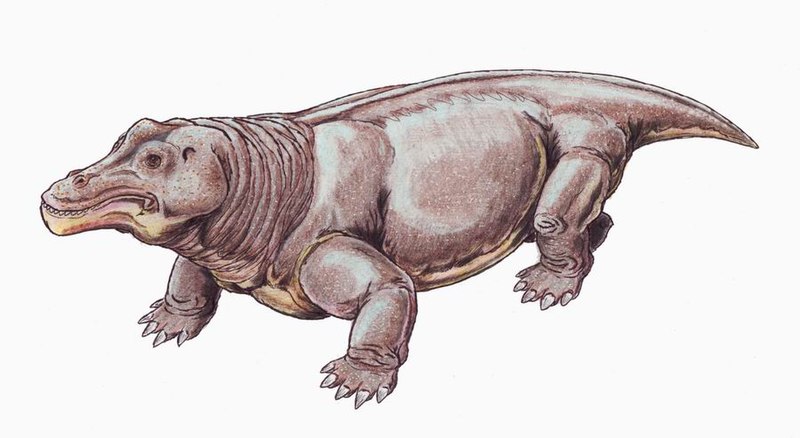
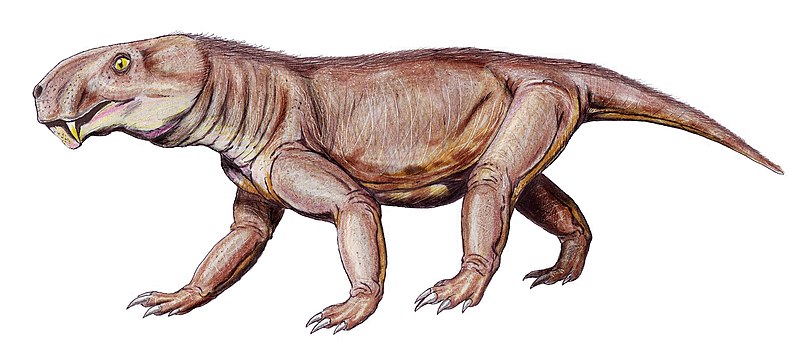
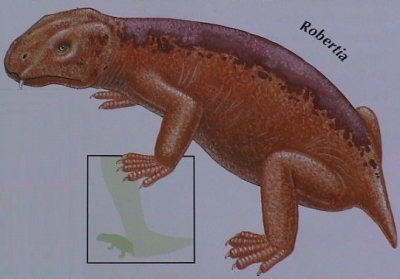
Robertia Робертия (Robertia Boonstra, 1948) — род ранних примитивных дицинодонтов из верхней перми Карру, Южная Африка (зона Tapinocephalus). Единственный вид рода — Robertia broomiana Boonstra, 1948. Robertia was a small, primitive dicynodont, and among the earlier members of this group. The type species R. broomiana was named in 1981. It is known from the Tapinocephalus Assemblage Zone of the South African Karoo, and was about 20 centimetres (8 in) in length. Robertia possessed a moderately wide skull roof, small postcanine teeth, and the palatine bone in the roof of its mouth was not as reduced as in its more successful relative, Diictodon. It had a notch immediately in front of the tusk-like canines on the upper jaw, which would presumably have held tough plant matter, such as stems and twigs, before the animal severed them with its horny beak.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 10:31 pm [industrialterro] [Link] |
Proburnetia Пробурнетия (Proburnetia viatkensis) — тероморф из позднепермских отложений Кировской области (Ильинский субкомплекс). Относится к семейству бурнетиид, сближаемому с эотериодонтами (биармозухиями) или с горгонопсами. Отличается развитием многочисленных выростов на черепе (надглазничные кости, носовые кости, заглазничные кости и т.д). Так называемые «рога» несут отпечатки сосудов и не могли быть покрыты роговыми чехлами. Назначение «рогов» остается неизвестным — существует предположение об их участии в выработке гормонов. Возможна защитная функция (защищали глаза в зарослях). Описана верхняя носовая раковина (турбиналь), хоаны сдвинуты вперед, костного вторичного неба нет, но мягкие ткани могли образовывать неполное вторичное небо. Длина черепа пробурнетии — 21 см, животное было размером с собаку. Вероятно, всеядное животное. Другие бурнетииды найдены в основном в Южной Африке. В 2006 году в Южной Африке в примерно одновозрастных с Ильинским комплексом отложениях найдена парабурнетия (Paraburnetia sneeubergensis), очень сходная с пробурнетией.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 10:26 pm [industrialterro] [Link] |
Porosteognathus Поростеогнат (Porosteognathus efremovi) – среднепермский тероцефал из Ишеевской фауны нижнетатарского подъяруса в Татарстане. Известен по фрагментам черепов, описан Б. П. Вьюшковым в 1952 г. Длина черепа достигала 16 см. Зубная формула обычно указывается как I6-C2-Pc9, что предполагает наличие двух верхних клыков. В действительности, второй клык был замещающим. Размеры костей в сборах предполагают наличие полового диморфизма. Является специализированным наземным хищником. Согласно М. Ф. Ивахненко (2001) из ишеевских сборов описаны теменные кости какого-то тероцефала, вероятнее всего – поростеогната. Эти остатки позволяют установить, что поростеогнат имел два параллельных костных гребня, проходивших по крыше черепа между орбитами очень близко друг к другу. Возможно, гребни служили «рассекателем» для защиты глаз при передвижении в зарослях. С другой сторны, гребни могли отводить тепло от головы. Есть определенное сходство между орнаментацией черепа бурнетиаморф и поростеогната. Часто поростеогната считают родственником южноафриканского пристерогната, но поростеогнат может принадлежать к особой ветви восточноевропейских тероцефалов.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 10:14 pm [industrialterro] [Link] |
Pristerognathus Пристерогнат (Pristerognathus) — примитивный тероцефал пермского периода. Происходит из «среднепермских» отложений Южной Африки, характеризует одноименную зону отложений формации Кароо. Ранее зона Pristerognathus считалась верхней частью зоны Tapinocephalus. Череп низкий, длинный, развиты крупные клыки, заклыковые зубы относительно немногочисленны. Высокий сагиттальный гребень. Морда высокая, узкая. Внешне, вероятно, несколько напоминал волка — с длинными мощными ногами и довольно коротким хвостом. Довольно крупный хищник, длина черепа до 30 см, общая длина более 1,5 метров. Типовой вид — P. polyodon, описан Г. Сили в 1895 году. Наиболее часто изображается крупный вид P. vanderbyli. Примерно 5 видов.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 10:03 pm [industrialterro] [Link] |
Pravoslavlevia Православлевия (Pravoslavlevia Vjuschkov, 1953) — род некрупных горгонопсов из верхней перми Восточной Европы. Название дано в честь профессора Санкт-Петербургского университета П. А. Православлева, ученика В. П. Амалицкого. Входит в состав северодвинской фауны (соколковский субкомплекс соколковского комплекса) вместе с иностранцевией.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:52 pm [industrialterro] [Link] |
Phthinosuchus Фтинозух (Phthinosuchus discors) — примитивный терапсид, обычно относимый к группе эотериодонтов (биармозухий), а в последнее время сближаемый с горгонопсами-рубиджеидами. Известен по неполным черепам из «средней» перми Приуралья (так называемые медистые песчаники). Впервые описан как «голова ящера» фон Кваленом в 1840 г. Фишер фон Вальдгейм первоначально отнес череп к роду Rhopalodon. Затем он описал другой образец под названием «Dinosaurus murchisoni» (в 1847 году). Название Dinosaurus отражало «хищную и прожорливую натуру» этого животного. Эйхвальд в 1848 году доказал принадлежность фрагментов одному черепу. Но он не считал возможным использовать название Dinosaurus, поскольку к тому времени Р. Оуэн уже предложил это название для целого отряда вымерших рептилий. Повторно описан Г. Сили под именем «Rhopalodon» в 1894 году, причем именно этот автор описал другой почти полный череп, изображаемый в научной литературе. Род Phthinosuchus предложен И. А. Ефремовым в 1954 году. Ефремов считал фтинозуха примитивным горгонопсом. Истинное систематическое положение данного рода оставалось неуточненным, ввиду фрагментарности остатков. В последнее время М. Ф. Ивахненко вновь отнес животное к горгонопсам, сближая его с рубиджеями. Ивахненко настаивает на приоритете названия Dinosaurus murchisoni и в последней (2008) сводке по пермским тетраподам фтинозух назван именно так. Длина черепа составляла около 20 см, череп высокий и узкий, крупная глазница с кольцом склеротики. Передняя часть черепа неизвестна. Около 10−12 пар сжатых с боков заклыковых зубов в верхней и нижней челюсти. В целом череп сильно деформирован и в настоящее время для изучения доступны лишь слепки исходных находок и череп, описанный Г. Сили. Часто публикуемая реконструкция черепа фтинозуха (и модель черепа в Палеонтологическом музее в Москве) основана на работе И. А. Ефремова, передняя часть черепа в этой реконструкции восстановлена. Несомненный хищник, длиной около 1,5 метров. Возраст фтинозуха, вероятно, соответствует Очерскому субкомплексу (около 267 млн л.н.), но может быть и несколько моложе. Близкий род — Камагоргон (Kamagorgon ulanovi) — описан Л. П. Татариновым в 1999 году из более древних отложений голюшерминского комплекса в Удмуртии.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:46 pm [industrialterro] [Link] |
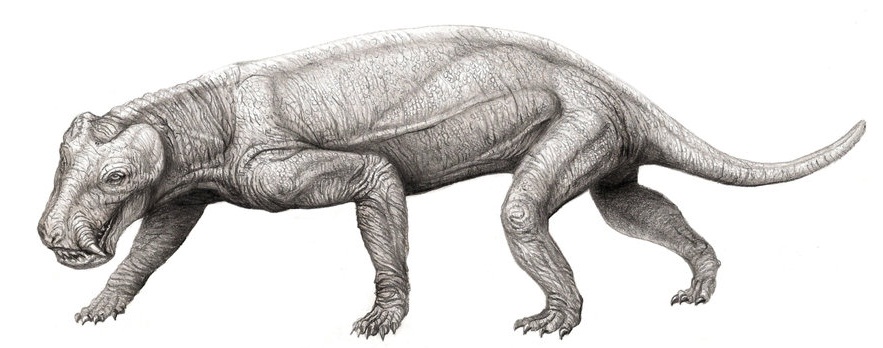
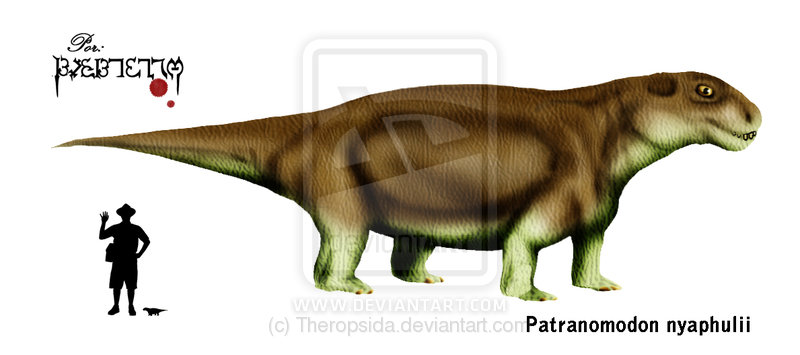
Patranomodon Patranomodon is an extinct genus of primitive anomodont. Patronomodon is primitive in that it retains a complete set of teeth in both jaws, in contrast to their descendants, the dicynodonts, in which the dentition is reduced to only a single pair of tusks (an in many cases no teeth at all), with their jaws covered by a horny beak similar to that of the modern tortoise although they are in no way closely related to each other. It was the size of a mongoose. Its discovery from the earliest terrestrial rocks of Gondwana (from Prince Albert in the southern Cape Province of South Africa) has shown that this group of herbivores originated in Gondwana rather than Laurasia, as had previously been supposed, and spread to other Pangaean continents. Аномодонты (Anomodontia) — подотряд терапсид, живший со средней перми по триас. Очень разнообразная группа, включающая в основном растительноядных животных от 20 см до 3,5 метров и более в длину. Одна из трех главных групп терапсид наряду с горгодонтами и териодонтами, возможно происходит независимо прямо от пеликозавров.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:30 pm [industrialterro] [Link] |
Procynosuchus Procynosuchus (Greek: "Before dog crocodile") was a cynodont from the Late Permian. It is considered to be one of the earliest and most basal cynodonts. Remains of Procynosuchus have been found in Germany, Zambia and South Africa. It was 60 cm (2 ft) long. As one of the earliest cynodonts, Procynosuchus has many primitive features, but it also has features that distinguish it from all other early therapsids. Some of these features have been interpreted as adaptations for a semi-aquatic lifestyle. For example, the wide zygapophyses of the vertebrae allow for a high degree of lateral flexibility, and Procynosuchus may have used anguilliform locomotion, or eel-like undulation, to swim through the water. The tail of Procynosuchus is also unusually long for a cynodont. The long haemal arches would have given the tail a large lateral surface area for greater propulsion through the water. Relatively flat foot bones may also have been an adaptation toward swimming, as the feet may have been used like paddles. Ridges on the femur are an indication of strong flexor muscles that could have stabilized the leg during limb-driven swimming. When the thigh is pulled back in the water, the lower leg tends to bend forward. Strong flexor muscles would have pulled the lower leg back with the femur, providing the powerful backward thrust that is needed to swim. Цинодо́нты (Cynodontia — «собачьи зубы») — вымершая группа териодонтов («зверозубых ящеров»), относящихся к отряду звероподобных рептилий — терапсид. Являлись предками млекопитающих и обладали почти всеми их признаками (больше всего напоминали современных однопроходных). Зубы были полностью дифференцированы (их дифференциация продолжалась в течение всего периода существования цинодонтов). Цинодонты откладывали яйца, подобно рептилиям. Скорее всего, они были теплокровными и покрытыми шерстью. Инфраотряд Процинозухии (Procynosuchia) — наиболее примитивные цинодонты, в основном позднепермские. Вторичное нёбо частично неокостеневшее. Щёчные зубы мелкие, многочисленные, трикуспидальные. Череп обычно низкий. Конечности мощные, пальцы длинные. Некоторые, возможно, полуводные. Два семейства: Двинииды (Dviniidae) — единственный род и вид (Двиния прима Dvinia prima) из поздней перми Восточной Европы (Архангельская область). Двинии — мелкие животные со специализированными щёчными зубами, возможно, растительноядные. Процинозухиды (Procynosuchidae) — из поздней перми Южной Африки (зона Dicynodon) и Европы. Всеядные, возможно, полуводные животные с длинным телом и широкими кистями и стопами. 5-6 родов, наиболее известен процинозух (Procynosuchus) из Южной Африки и Германии.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 09:22 pm [industrialterro] [Link] |
Oudenodon Oudenodon is an extinct genus of dicynodont. It was common throughout southern Africa during the Late Permian. Several species of Oudenodon are known. Both O. bainii, the type species, and O. grandis are known from South Africa. Specimens of O. luangwensis have been found from Zambia. One species, O. sakamenensis, is the only therapsid yet known from Madagascar. It is the type genus of the family Oudenodontidae, which includes members such as Cteniosaurus, Tropidostoma, and Rhachiocephalus.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| November 13th, 2011 | |
| 05:02 pm [industrialterro] [Link] |
Nikkasaurus Никказавр (Nikkasaurus) — представитель примитивных терапсид из «средней» перми России. Очень мелкое животное (длина черепа около 5 см). Морда заостренная, низкая. Глазницы огромные, с кольцом склеротики. Височное окно гораздо меньше глазницы. Затылок наклонен назад, как у всех терапсид. Тем не менее, внешне череп поверхностно сходен с черепом пеликозавров (в частности, варанопсеид). Зубы мелкие, «клыки» не выделяются. Передние зубы игловидные, задние зубы расширеные, трехвершинные. Скелет легко построен. Вероятно, питались преимущественно насекомыми, могли быть ночными животными. Единственный вид (N. tatarinovi) описан М. Ф. Ивахненко в 2000 году из мезенского субкомплекса Очерского комплекса «средней» перми. Эти отложения в Архангельской области соответствуют позднеказанскому (капитаний) веку пермского периода. Систематическое положение никказавра остается неясным — вероятно, он относится к терапсидам, но может быть и представителем более примитивных синапсид. К семейству Nikkasauridae относят также род Reiszia с двумя видами (R. gubini и R. tippula), описанными Ивахненко также в 2000 году из этих же отложений. Ивахненко сближал никказавров с примитивными тероцефалами. Никказавры могут быть реликтами более древних этапов развития терапсид. В последней (2008) сводке по пермским терапсидам М.Ф. Ивахненко относит никказавров к основанию подотряда Gorgonopsia, в качестве одноименного инфраотряда Nikkasaurida.
Tags: Вымершие синапсиды, Пермь, терапсиды |




























































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}