[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 20 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| January 31st, 2012 | |
|---|---|
| 07:18 pm [industrialterro] [Link] |
Exaeretodon Exaeretodon is a genus of traversodontid cynodont; several species are known, from various formations. E. argentinus, E. frenguelli, and E. vincei are from the Carnian-age (Upper Triassic) Ischigualasto Formation of Argentina. E. major and E. riograndensis are from the Ladinian-age (Middle Triassic) portion of the Santa Maria Formation of Brazil. E. statisticae is from the Carnian-age Lower Maleri Formation of India. This genus was an herbivore up to 1.8 meters long (5.9 ft), with a specialized grinding action when feeding. An analysis of the size of the bones of calves collected in Paleorrota concluded that the mother Exaeretodon had one or two calves, for one pregnancy. Another point of interest is that these cynodonts had deciduous teeth, which is a characteristic of mammals and means that babies could not chew, and required specialized parental care. Only older juveniles had permanent teeth. Exaeretodon is one of the largest traversodontids, but the traversodontid species Ischignathus sudamericanus was reported in 1967 to be even larger. I. sudamericanus was based on a single skull from the Ischigualasto Formation in the same deposit as Exaeretodon argentinus. When he first named the species, Argentine paleontologist José Bonaparte mentioned several features that distinguish it from all other traversodontids. The tooth rows of the upper jaw are more parallel to each other in I. sudamericanus than they are in Exaeretodon, and they are also inset closer to the inside of the mouth. There are also more postcanine teeth oriented toward middle of the subtemporal fenestrae (two holes in the bottom of the skull) and ascending rama of the dentary (projections of the lower jaw that extend up to the skull). The ascending rama are also wider and taller in the I. sudamericanus specimen. The orbit or eye socket is longer than that of Exaeretodon, as are the palatine bones. Despite the differences, a 2007 study concluded that the I. sudamericanus skull represents the same species as E. argentinus. Using allometry, paleontologist Jun Liu found I. sudamericanus to be the largest known example of a growth series in E. argentinus. Thus, Ischignathus is now regarded as a junior synonym of Exaeretodon. As the animal grew, the proportions of bones changed. These differing proportions were initially seen as species-distinguishing characters, but are now regarded as natural ontogenic variation.
Tags: Вымершие синапсиды, Триас, терапсиды |
| 06:51 pm [industrialterro] [Link] |
Euchambersia Эучамберсия или эвчамберсия (Euchambersia mirabilis) — хищный тероцефал позднепермской эпохи, близкий к мосхорину, териогнату и аннатерапсиду. Предположительно, имела ядовитые железы, соединявшиеся с бороздами на верхних клыках. Череп высокий, заклыковые зубы отсутствуют, пять резцов и один клык с каждой стороны. Теменного отверстия нет. Отсутствует (вероятно) также полная скуловая и заглазничная дуги. Нёбные отверстия малы. Позади верхних клыков — глубокие вырезки, возможно, вмещавшие ядовитую железу, от этой полости к клыку идет ложбинка. Вырезка соединяется с двумя крупными отверстиями под глазницей (возможный выход сосудов и нервов). На самом клыке есть бороздка и гребень, для стока секрета ядовитой железы, причем борозда идет по передней поверхности клыка. Вторичное небо не развито. Длина черепа 10 см. Скелет и нижняя челюсть неизвестны. Обнаружена в позднепермских отложениях (зона Cistecephalus) Кароо в Южной Африке. Вероятно, питалась относительно некрупной добычей, которую могла проглотить целиком. Тем не менее, не исключено также питание относительно крупными животными типа дицинодонтов. В таком случае, вероятно, эучамберсия наносила добыче укус, а затем ожидала ее гибели, преследуя умирающее животное. Такая тактика, тем не менее, могла быть не очень удачной в условиях конкуренции с другими хищниками (какой-нибудь горгонопс или другой тероцефал мог перехватить ослабевшую добычу).
Tags: Вымершие синапсиды, Триас, терапсиды |
| 06:35 pm [industrialterro] [Link] |
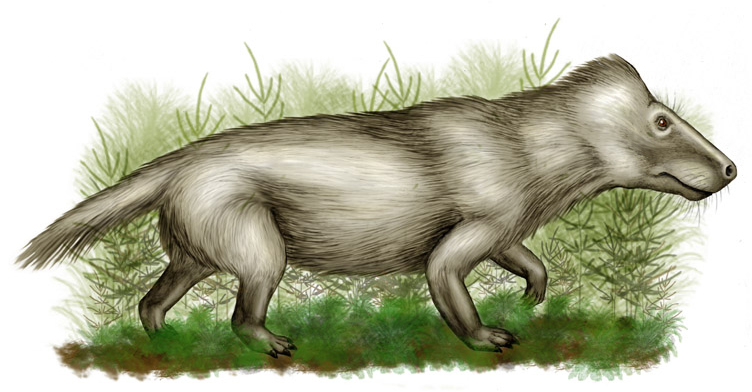
Ericiolacerta Эрициолацерта (Ericiolacerta) — мелкий тероцефал раннетриасовой эпохи, принадлежащий к бауриаморфам. Описана Д. Уотсоном в 1931 году. Единственный вид - E. parva из зоны Lystrosaurus раннего триаса Южной Африки. Мелкое животное, общая длина до 20-25 см. Череп грушевидный, удлиненный, уплощенный. Заглазничная дуга неполная. Теменного отверстия нет. Хорошо развито вторичное небо, образованное сошником и небными костями. Квадратные ветви крыловидных костей очень тонкие. Предчелюстная и челюстная кость покрыты множеством отверстий (трактуются как сосудистые отверстия, указывающие на развитие мягких тканей лицевой области). Нижняя челюсть длинная и тонкая, с зачаточным венечным отростком. Симфиз нижней челюсти длинный и скошенный. Зубы мелкие, цилиндрические, клыки не выделяются, первая пара нижнечелюстных зубов направлена косо вперед (“резцы”). Заклыковые зубы со слабо трехраздельными тупыми коронками. Скелет довольно компактный, выражен поясничный отдел позвоночника, три крестцовых позвонка. Хвост короткий. Лопатка длинная и тонкая, верхний конец ее приближен к черепу. Таз низкий, короткий, широкий, с тироидными отверстиями. Кости конечностей длинные, тонкие. Стопа пальцеходящая. В целом по очертаниям скелета животное несколько напоминало ежа. По-видимому, этот высокоразвитый тероцефал был покрыт шерстью, имел развитые вибриссы на морде и внешне походил на млекопитающее. Питалась эрициолацерта насекомыми и другими беспозвоночными, занимая экологическую нишу современных насекомоядных. Остатки этого вида известны и из одновозрастной формации Фремоу в Антарктиде.
Tags: Вымершие синапсиды, Триас, терапсиды |
| 06:19 pm [industrialterro] [Link] |
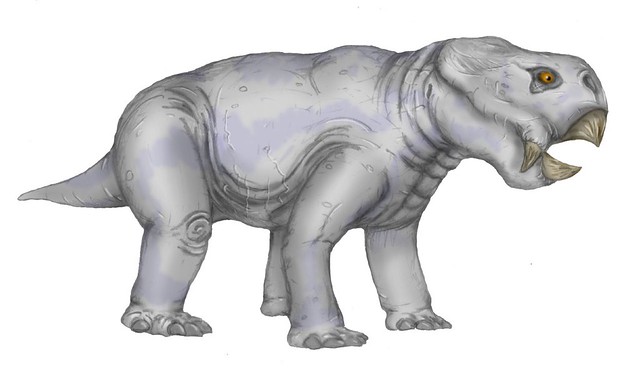
Dinodontosaurus Dinodontosaurus (meaning "terrible-toothed lizard") is a genus of dicynodont therapsid. It was one of the largest herbivores of the Triassic (about 8 feet (2.4 m) long and weighing a few hundred pounds) and had a beak corneum. It lived in the Middle Triassic but disappeared in the Upper Triassic. Dinodontosaurus turpior is the most common species of dicynodont that existed in the Middle Triassic, and more common in the fossil layers that age in Rio Grande do Sul, in geopark of paleorrota. They are found mainly in the Paleontological Site Chiniquá in São Pedro do Sul and Candelária , where a group of ten pups were found together, demonstrating that these animals had strategies for coexistence in a group and caring for their offspring. Dinodontosaurus pedroanum was a specie of dicynodonts herbivore that lived in the region of paleorrota, Brazil (8 fossils collected) and Argentina (2 fossils collected). It was described the first time by Tupi Caldas in 1936 and is synonymous Dinodontosaurus oliveirai, Romer 1943. Каннемейериды (Kannemeyeridae seu Kannemeyeriiformes) — семейство крупных дицинодонтов триасовой эпохи. Возможно, потомки листрозавров. Доминирующие растительноядные в течение большей части триаса. Дицинодонт из раннего мела Австралии также, вероятно, относится к этой группе. Группа довольно разнообразна, различные её представители отличаются не только размерами (от овцы до носорога) но и формой и пропорциями черепа и конечностей. Динодонтозавры (Dinodontosaurinae) — единственный род Dinodontosaurus из среднего триаса Южной Америки, 3 вида. Длинноногие короткотелые дицинодонты с длинными верхними клыками, морда короткая.
Ископаемые останки (1, 2, 3, 4):
( Далее ) Tags: Вымершие синапсиды, Триас, терапсиды |
| 06:06 pm [industrialterro] [Link] |
Diademodon Diademodon is an vegetarian extinct genus of therapsid. It was about the size of a small cow. Fossils of the Diademodon tetragonus were found in levels of the Rio Seco de la Quebrada geological formation in Mendoza Province, Argentina.
Tags: Вымершие синапсиды, Триас, терапсиды |
| January 29th, 2012 | |
| 01:00 pm [industrialterro] [Link] |
Cynognathus Циногнат — ископаемый хищник длиной тела около метра, живший в период раннего и среднего триаса. Он принадлежал к числу синапсид, животных, более напоминающих современных млекопитающих, и входил в инфраотряд Eucynodontia. Род Cynognathus имел распространение почти по всему миру. Ископаемые останки циногнатов на сегодняшний день обнаружены на территории Южной Африки, Южной Америки, Китая и Антарктиды. Было описано много видов, но все они являются синонимами вида Cynognathus crateronotus. Cynognathus crateronotus was a meter-long predator of the Early to Middle Triassic. It was among the more mammal-like of the Synapsids, a member of a grouping called Eucynodontia. The genus Cynognathus had an almost worldwide distribution. Fossils have so far been recovered from South Africa, South America, China and Antarctica. The genus Cynognathus (from Greek κυνόγναθος, meaning "dog jaw") has been given several different names over the years. It has also been known as Cistecynodon, Cynidiognathus, Cynogomphius, Karoomys, Lycaenognathus, Lycochampsa, Lycognathus, and Nythosaurus. In addition, according to the records of the Peabody Museum of Natural History at Yale, Richard Owen used the name Nythosaurus for this animal in 1876. This usage seems to be unconnected with Cynognathus. Cynognathus is presently the only recognized member of family Cynognathidae. Opinions vary as to whether all remains belong to the same species. The species Cynognathus crateronotus is also known as Cistecynodon parvus, Cynidiognathus broomi, Cynidiognathus longiceps, Cynidiognathus merenskyi, Cynognathus beeryi, Cynognathus minor, Cynognathus platyceps, Cynogomphius berryi, Karoomys browni, Lycaenognathus platyceps, Lycochampsa ferox, Lycognathus ferox, Nythosaurus browni. Fifteen different names for one Mesozoic creature might be regarded as excessive, but it's by no means a record. The dinosaur Plateosaurus engelhardti, has been named well over 20 times. The genera Karoomys, Cistecynodon and Nythosaurus are known only from tiny juveniles, while Lycognathus cucullatus seems to be a misidentified snake from the Balearic Islands, although confirmation is elusive. Cynognathus was a heavily built animal, and measured around 1 metre (3.3 ft) in body length. It had a particularly large head, 30 centimetres (1.0 ft) in length, with wide jaws and sharp teeth. Its hind limbs were placed directly beneath the body, as in mammals, but the fore-limbs sprawled outwards in a reptilian fashion. The dentary was equipped with differentiated teeth that show this animal could effectively process its food before swallowing. The presence of a secondary palate in the mouth indicates that Cynognathus would have been able to breathe and swallow simultaneously. The lack of ribs in the stomach region suggests the presence of an efficient diaphragm: an important muscle for mammalian breathing. Pits and canals on the bone of the snout indicate concentrations of nerves and blood vessels. In mammals, such structures allow hairs (whiskers) to be used as sensory organs.
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие синапсиды, Триас, терапсиды |
| 12:50 pm [industrialterro] [Link] |
Chiniquodon Chiniquodon is a genus of carnivorous "mammal-like reptile", (Therapsid), which lived during the lower Upper Triassic in South America. These animals were up to the size of a dog. This genus seems to be most closely related to a contemporary genus, Probelesodon. In various respects, the anatomy is very mammal-like, though not strongly enough to be viewed as a direct ancestor.
Tags: Вымершие синапсиды, Триас, терапсиды |
| 12:42 pm [industrialterro] [Link] |
Boreogomphodon Boreogomphodon is a genus of traversodontid cynodont from the Late Triassic of the eastern United States. Fossils have been found from the Turkey Branch Formation in Virginia. Boreogomphodon jeffersoni was named in 1990. Its teeth and cranial bones are the most common tetrapod fossils in the Turkey Branch Formation. Boreogomphodon was distinguished from other traversodontids like the African Luangwa and the South American Traversodon on the basis of its postcanine teeth. Most traversodontids have lower postcanine teeth with two cusps, but Boreogomphodon was the first traversodontid found with three cusps on its lower postcanine teeth. There is a single cusp on the side of the upper postcanine facing the lip, while a flat surface extends outward from it. The traversodontid Arctotraversodon from the Wolfville Formation of Nova Scotia is similar to Boreogomphodon in that it has three cusps on its lower postcanine, but its postcanines are much wider than they are long relative to those of Boreogomphodon. Boreogomphodon specimens likely represent juvenile individuals because of their small size. The snout is short, while larger, presumably mature traversodonts have longer snouts. Boreogomphodon specimens have only four or five postcanine teeth. Traversodontids acquire more of these teeth as they grow, suggesting that Boreogomphodon individuals were not fully mature. Boreogomphodon also has some sectorial, or cutting, postcanine teeth, which are not present in most larger traversodontids. Boreogomphodon has several primitive features, including a depression in the upper jaw called the paracanine fossa in front of the canine tooth. This fossa provides space for the lower canine tooth when the jaw is closed. Above the upper canine at the top of the snout there is a small depression on the maxilla. The tip of the snout is covered in irregular pits, a unique feature of Boreogomphodon. Each upper postcanine has large central cusp and a posterior cingulum.
Tags: Вымершие синапсиды, Триас, терапсиды |
| 12:31 pm [industrialterro] [Link] |
Belesodon Белезодон (Belesodon) — род вымерших цинодонтов, известный по единственному виду Belesodon magnificus. Belesodon magnificus был первым из цинодонтов, обнаруженных в формации Санта-Мария в Палеорроте, Бразилия. Его впервые описал Фридрих фон Хюне в 1935—1942 гг. В 2002 г. Джанини и Абдала дали виду синонимичное название Chiniquodon theotonicu.
Tags: Вымершие синапсиды, Триас, терапсиды |
| 12:19 pm [industrialterro] [Link] |
Bauria Баурия (Bauria) — вымерший род подотряда тероцефалов семейства Bauriidae. Жил в раннем триасовом периоде. По образу жизни баурия была, вероятно, насекомоядной или хищником. Bauriidae is a family of therocephalian therapsids. Bauriids are among the most advanced eutherocephalians and possess several mammal-like features such as a secondary palate. Unlike other therocephalians, bauriids were herbivorous. Two subfamilies are classified within Bauriidae: Nothogomphodontinae and Bauriinae. Bauriids have a dentition characteristic of herbivores. There are four incisors on either side of the upper jaw. Like other therocephalians, bauriids have moderately enlarged canines. The postcanine teeth behind the canines are broad. The postcanines of the upper and lower jaw fit tightly together as an adaptation to processing plant material.
Tags: Вымершие синапсиды, Триас, терапсиды |
| December 5th, 2011 | |
| 06:04 pm [industrialterro] [Link] |
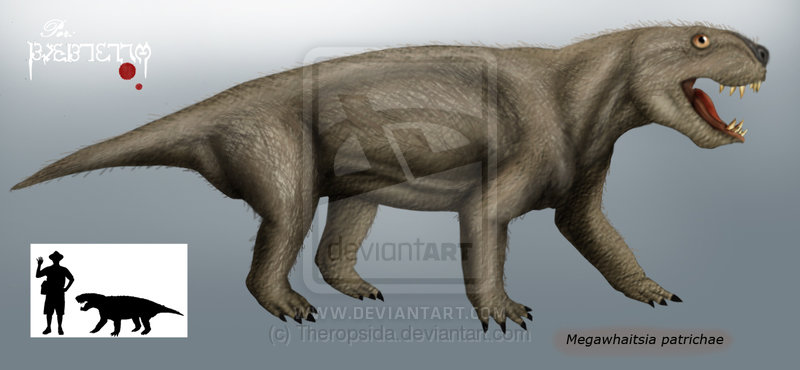
Megawhaitsia Мегавайтсия (Megawhaitsia patrichae) — гигантский тероцефал из терминальной поздней перми Владимирской области. Происходит из местонахождения Вязники-2, характеризующего наиболее поздние горизонты поздней перми. Известен по обломку правой верхнечелюстной кости. Первоначально эти остатки считали остатками горгонопса, но позже выяснилось, что в этой фауне горгонопсов уже нет. Скуловая и заглазничная кости полные, есть крупное отверстие для нижнечелюстного клыка в верхнечелюстной кости, за ним — альвеолы для трех крупных клыков (из них два первых несколько крупнее). Максилла очень массивная, корни клыков глубокие. Все три клыка имеют общую альвеолярную сумку. Под поверхностью кости проходят три канала, лежащие латеральнее альвеол клыков. Начало этих каналов лежит где-то в районе слезно-носового протока. Каналы открываются в районе альвеол каждого клыка. Длина фрагмента верхнечелюстной кости более 10 см, общая длина черепа была до 40-50 см. Вид упомянут (как неназванный мосхорининд) М. Ф. Ивахненко в 1997 году. Назван этим же автором в 2008 году. Видовое название дано в честь американского палеонтолога Патриции Викерс-Рич. Относится к семейству Whaitsiidae, куда принадлежит также южноафриканский териогнат. От африканских родственников мегавайтсия отличается огромными размерами. По-видимому, это объясняется отсутствием в вязниковском комплексе горгонопсий при сохранении фауны крупных дицинодонтов. Вакантное место крупного хищника было занято крупными протерозухиями Archosaurus — пришельцами с сухих возвышенностей — и гигантскими тероцефалами. Последние происходят от исконных обитателей побережий водоемов, мелких всеядных тероцефалов более ранних горизонтов перми. Мегавайтсия стала специализированным охотником на дицинодонтов. Это первый настоящий вайтсиид из Восточной Европы. Самым необычным анатомическим признаком мегавайтсии являются каналы в толще верхней челюсти. Ивахненко расценивает их как протоки ядовитых желез, примерно как у ящериц-ядозубов.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 05:56 pm [industrialterro] [Link] |
Kamagorgon Камагоргон (Kamagorgon ulanovi) — примитивный хищный терапсид среднепермской эпохи. Ближайший родич фтинозуха, включается в семейство фтинозухид. Возможно, относится к примитивным горгонопсам. Довольно крупное животное, череп до 40 −45 см длиной. Длинные уплощенные верхние клыки, предклыковые и заклыковые зубы режущие, массивные. Хорошо развит высокий носо-глоточный желоб, ограниченный с боков широкими буграми с мелкими небными зубами. Развит пахиостоз лобных костей, максиллы скульптированы мелкими ямками. Известна только передняя часть черепа. Крупный наземный хищник, охотился на растительноядных парарептилий и терапсид. Единственный вид — Kamagorgon ulanovi, описан Л. П. Татариновым в 1999 г из отложений казанского яруса в Удмуртии (Сидоровы Горы). Данные отложения относятся к голюшерминскому субкомплексу, более древнему, чем Очерская фауна. Видовое название дано в честь местного геолога Е. И. Уланова, нашедшего череп. Описанный И. А. Ефремовым в 1954 году Admetophoneus kargalensis из средней перми Каргалинских рудников в Оренбургской области может быть близок к камагоргону. Этого гигантского (с черепом около 55 см длиной) хищника Ефремов отнес к антеозаврам. Адметофонеус описан по фрагменту верхнечелюстной кости и истинное его положение в системе неизвестно.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 05:43 pm [industrialterro] [Link] |
Viatkogorgon Вяткогоргон (Viatkogorgon ivachnenkoi) – мелкий горгонопс из так называемой Котельнической фауны (середина поздней перми, фауна обнаружена на берегах реки Вятки в Кировской области), примерно соответствующей по возрасту африканской зоне Tropidostoma. Полный скелет найден в 1992 г. Род вяткогоргон описан академиком Л. П. Татариновым в 1999 г. Включает один вид – Viatkogorgon ivakhnenkoi Tatarinov, 1999 Вяткогоргон был подвижным наземным или полуводным плавающим хищником, возможно, факультативным падалеядом. Его скелет найден рядом со скелетом дромазавра суминии (Suminia getmanovi). Вяткогоргон мог охотится на суминий, дицинодонтов, тероцефалов, молодых парейазавров дельтавятий (Deltavjatia vjatkensis), также входивших в состав котельнической фауны.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 05:27 pm [industrialterro] [Link] |
Sinophoneus Синофонеус (Sinophoneus yumenensis) – представитель хищных дейноцефалов из «средней» перми Китая. Принадлежит к семейству Anteosauridae, известному также из Южной Африки и Приуралья. Особенно близок к роду титанофонеус из чуть более поздних отложений Татарии. Почти полный череп синофонеуса был обнаружен в 1981 году в богатом пермском местонахождении Дашанкоу в западном Шанси. Это первая находка дейноцефала в Восточной Азии. Род и вид описаны в 1995 году. Название рода переводится как «китайский убийца» и отражает родство с титанофонеусом. Длина черепа 35 см, затылочная часть относительно широкая. Ноздри небольшие, на кончике морды. Орбиты направлены вперед и в стороны, овальные, относительно небольшие. Пахиостоз дорзального края орбиты слабый. Предлобная часть крыши черепа слабо пахиостозна, с гребнями, отделяющими дорзальную поверхность от боковых. Умеренно выражен срединный гребень в предлобно-теменной части крыши черепа. Теменное отверстие небольшое, расположено на возвышении, как обычно для антеозавров. Скуловая дуга высокая кзади. Хоаны длинные и глубокие, достигают передним краем уровня клыков. Очень массивные птеригоиды. Зубная формула верхней челюсти – 5 резцов, 1 клык, 8-9 заклыковых зубов. Клыки очень крупные, последний резец небольшой. Заклыковые зубы по размерам сравнимы с резцами, образуют один однородный ряд. Хищное животное, в рационе питания которого большое значение могла играть рыба и низшие тетраподы. Слои Дашанкоу традиционно считались верхнеказанскими – нижнетатарскими по возрасту, аналогично фауне Ишеево в Татарстане. Недавние исследования показали, что китайская фауна может быть более древней, относящейся к уфимскому (роадскому) времени. В таком случае синофонеус является древнейшим из дейноцефалов, наряду с микросиодоном из фауны Голюшермы в Приуралье.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| December 4th, 2011 | |
| 04:22 pm [industrialterro] [Link] |
Venjukovia Venjukovia (Amalitskii, 1922) is an extinct genus of non-mammalian synapsid from the Middle Permian of Russia. Венюковии (Venyukoviidae) — семейство примитивных аномодонтов среднепермской эпохи. Образуют надсемейство Venyukovioidea. Описаны два рода, которые могут быть синонимами. Венюковия (Venyukovia prima) описана В. П. Амалицким по нижним челюстям из Каргалинских рудников в Оренбуржье. Остатки собрал в 1908 году профессор Петроградского горного института П. Н. Венюков, в честь которого и был назван род. Владимир Амалицкий считал венюковию млекопитающим. Позднее, в 1935 году череп подобного животного был обнаружен в Ишеево. Изучение ишеевского черепа и челюстей венюковии позволило И. А. Ефремову отнести животное к терапсидам. Ефремов сближал венюковию с дейноцефалами. Позднейшие исследования позволили установить её принадлежность к самым примитивным аномодонтам. Для черепа из Ишеева обнаружили отличия в строении зубов и он был отнесён к особому роду улемика (Ulemica). Таким образом, от венюковии известны только обломки нижних челюстей. В 1960 году П. К. Чудинов описал из Очерского местонахождения череп мелкого аномодонта очерии (Otsheria netzvetajevi). Это ближайший родич венюковии, который может относиться к этому же роду. В целом находки этой группы очень редки. Также к семейству венюковий отнесена суминия, обнаруженная близ Котельнича. Венюковии — некрупные животные, длина черепа 10—12 см. Череп довольно высокий, короткий, полукруглый, глазницы огромные. Передние зубы долотовидные, напоминают резцы грызунов. Щёчные зубы широкие, сильно стёртые. Возможно, могли перерабатывать пищу во рту (раздавливать). Хоанная область очень глубокая. Теменной бугор высокий. Нижняя челюсть массивная. Скуловая дуга изогнутая, по очертаниям череп сходен с таковым у дицинодонтов. Вероятно, наземные растительноядные, питались относительно мягкой растительностью. Могли питаться также насекомыми.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 04:05 pm [industrialterro] [Link] |
Ulemosaurus Улемозавр (Ulemosaurus) — растительноядный дейноцефал из подотряда тапиноцефалов. Обитал в среднепермскую (около 265 млн л.н.) эпоху в Приуралье. Описан А. Н. Рябининым в 1938 году из района села Ишеево в Татарии. В 1980-х годах считался видом африканского рода мосхопс. Тем не менее, улемозавр — более примитивный тапиноцефал, иногда выделяемый в особое семейство. Типовой вид рода - Ulemosaurus svijagensis. Череп высокий, верхние края глазниц утолщены и бугристы, длина черепа свыше 40 см. Височная впадина узкая. Передние зубы (резцы) мощные, уменьшаются кзади. На зубах сильно развит уступ («пятка»), зубы могут плотно смыкаться. Сохраняются небольшие клыки, почти неотличимые от других зубов. Предполагается, что зубная система была приспособлена к перетиранию растительного материала, но перетирание осуществлялось передними зубами, а не задними, как у большинства растительноядных позвоночных. Улемозавр (и все прочие тапиноцефалы) мог питаться мертвыми гниющими стволами каламитов и других растений. Передние ноги длиннее и массивнее задних, голова высоко приподнята над землей (как и у всех тапиноцефалов). Плечевой пояс заметно массивнее, чем у мосхопса, плечевая кость длиннее. Общая длина животного могла быть до 4 метров. Описан (по фрагментам) еще один вид — Ulemosaurus gigas. Он известен по отдельному резцу из Ключевского рудника (Башкирия) и частичному черепу из местонахождения Озерки (Оренбургская область). Судя по всему, кроме более крупных размеров, отличался от типового вида более длинной мордой.
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 03:58 pm [industrialterro] [Link] |
Ulemica Улемика (Ulemica) — примитивный аномодонт среднепермской эпохи. Близка к венюковиям. Первоначально описана И. А. Ефремовым в 1940 году по черепу из Ишеевского местонахождения как второй вид венюковии (Venyukovia invisa). Долгое время череп изображали в литературе как череп венюковии. Исследования М. Ф. Ивахненко позволили в 1996 году выделить это животное в особый род и в отдельное семейство. Более полный череп из Оренбургской области, изображённый, в частности, в работе П. К. Чудинова как череп типового вида венюковии, принадлежит особому виду улемики — U. efremovi. Длина черепа улемики около 15 см, скелет неизвестен. Остатки встречаются гораздо чаще, чем остатки венюковий — до 15 % находок в Ишееве. Череп по очертаниям напоминает череп дицинодонтов. Нижняя челюсть очень массивная, с лопастевидными выростами на подбородке. Передние резцы мощные, как у грызунов, щёчные зубы сильно расширены. Очень широкая и мощная поверхность предчелюстных, челюстных и зубных костей предполагает ороговение. Посередине верхней челюсти есть один увеличенный зуб, входящий в ямку на нижней челюсти. Боковые движения челюстей при таком строении невозможны, челюсти могли лишь раздавливать пищу. Ефремов считал, что «венюковия» (улемика) питалась подобно грызунам, добывая пищу из твёрдых оболочек. На самом деле, улемики, вероятно, питались твёрдой пищей, которую раздавливали зубами. Пищей могли быть стволики молодых каламитов.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 03:43 pm [industrialterro] [Link] |
Titanosuchus Titanosuchus ferox ("Fierce Titan crocodile") was a dinocephalian therapsid that lived in the Mid Permian epoch in South Africa. Despite the name, it was not related to crocodiles. Along with its close relatives, Jonkeria and Moschops, Titanosuchus inhabited present-day South Africa around 255 million years ago, in the Late Permian. Titanosuchus was a carnivore and might have eaten both Jonkeria and Moschops, among other vertebrates. Its teeth included sharp incisors and fang-like canines, perfect for biting prey. Titanosuchus rivals that to Titanophoneus, which is also a carnivore and a dinocephalian, but it lived only in Russia. Titanosuchus should not be confused with the therapsid Eotitanosuchus, which belonged to a different family.
Tags: Вымершие синапсиды, Пермь, терапсиды |
| 03:09 pm [industrialterro] [Link] |
Titanophoneus Титанофонеус (Titanophoneus) — хищный дейноцефал группы антеозавров. Известен из «средней» перми Приуралья и Поволжья (описан из Ишеево в Татарии — нижнетатарский ярус). Во многом сходен с южноафриканским антеозавром. Череп высокий и узкий, у взрослых особей развиты утолщения заглазничной и лобной области (пахиостоз). Мощные резцы и клыки, 8—9 пар мелких заклыковых зубов. Ноги мощные, не очень длинные, с широкими кистями и стопами (возможно, имелась плавательная перепонка). Хвост длинный. В целом скелет довольно легкий. Молодые особи (первые из описанных экземпляров) достигали в длину 3 метров, черепа взрослых могли достигать 60 см в длину. Типовой вид — T. potens, описан И. А. Ефремовым в 1938 году. Типовой череп (молодая особь) сломан и его передняя часть резко вздернута вверх (именно так он изображался в старых книгах). В реальности он был не столь «бульдогообразным». К данному роду и виду может принадлежать и другой хищный дейноцефал из Ишеево — долиозавриск. М. Ф. Ивахненко обосновывает синонимичность типового вида титанофонеуса и долиозавриска (на чем изначально настаивал И. А. Ефремов). Ископаемые остатки титанофонеев обнаружены в ряде местонахождений Приуралья и Поволжья и датируются серединой поздней перми (уржумский горизонт, нижнетатарский подъярус, уржумский ярус, биармийский отдел). Остатки встречаются довольно часто — к примеру, в коллекции Ишеево они составляют до 15% всех находок, в том числе отсюда известно 2 более или менее полных скелета. Титанофонеи являются одними из лучше всего изученных представителей группы. Они представляли собой крупных дейноцефалов с большой головой, короткой мощной шеей, удлиненным приземистым туловищем, расставленными в стороны сильными конечностями и длинным хвостом. Черепа взрослых животных могли достигать 60 см в длину при общей длине тела около 5 м. В своих фаунистических сообществах они были наиболее крупными хищниками. Череп умеренно высокий, суженный спереди и расширенный в скуловой области, треугольная в сечении лицевая часть несколько удлинена (посторбитальное расстояние укладывается в общую длину черепа 4,5-5 раз). Край верхней челюсти впереди клыков полого поднимается вверх, не образуя резкого уступа. Подобная «бульдогообразность» присутствовала уже у раннепермских пеликозавров и, очевидно, связана со способом хватания добычи. Ноздри и орбиты несколько приподнятые. Глазницы сравнительно небольшие, округлые; глазные впадины направлены косо в стороны и довольно сильно вперед, что было важно при высматривании добычи. Глазное яблоко снабжено кольцом склеротики. Глазничный край скуловой кости с возрастом утолщался. Предлобная кость и верхний край глазницы с бугристой поверхностью. Межглазничный и теменной отделы сужены. Височная дуга высокая, утолщающаяся с возрастом. Задневисочные окна крупные, ямы большие, широко открытые сверху; задний край глубоко вклинен в отросток скуловой кости. Имеется редуцированная надвисочная кость, у старых животных сливающаяся с соседними костями черепа. Межтеменная кость очень высокая, сильно расширенная внизу, с мощным сагиттальным гребнем. Строение задней части черепа очень характерно для антеозавроидей, с отверстием теменного глаза, лежащем на заднем краю на высоком бугре, образованном одними теменными костями. Теменное отверстие крупное, слабо удлиненно-овальное, окруженное высоким костным воротничком; нерв теменного глаза был довольно длинным. Столбик среднего уха (стремечко) имеет вид небольшой распорки в основании черепа между квадратной костью и мозговой коробкой, где он упирается в овальное окно. Это положение столбика как будто бы исключает его подвижность и приспособленность для передачи звуковых колебаний через овальное окно во внутреннее ухо, хотя по своему местоположению он и равнозначен стремечку среднего уха млекопитающих, птиц и пресмыкающихся, имеющих барабанную перепонку. Как воспринимали звук титанофонеи — до конца неясно, но вряд ли животные были глухими. На задней половине носовых костей, предлобных, лобных и заднелобных костях с возрастом развиваются мощные пахиостозные образования с грубой ругозной поверхностью. Пахиостозный бугор назофронтальной области невысокий, захватывает значительную часть носовой кости, полого поднимаясь почти от премаксиллы. Ругозистые утолщения на заглазничной кости высокие, гребневидные. Вся область покрыта сетью борозд кровеносных сосудов. Наличие структур такого типа довольно обычно для растительноядов (например, относительно близких тапиноцефалоидей), но вызывает удивление у хищников — особенно в данном случае, учитывая вытянутую форму черепа. Частично их можно было бы толковать как итог приспособления к большой механической нагрузке со стороны переднего конца морды при хватании и удержании крупной добычи. Однако, у южноафриканских дейноцефалов подобные утолщения намного сильнее и, главное, слишком велики, чтобы в них можно было видеть отражение механической нагрузки. Не объясняет данная версия и странного бугристого строения внешней поверхности костей верхней стороны черепа. Зубная формула: I5/4, С1/1, РС6-8/6-9. Предклыковые зубы крупные, конические, с язычными гребнями и слабыми пятками, в которые упирались вершины зубов противоположной челюсти. В ряде случаев резцы скошены вперед, особенно верхние, очень плотно входящие в промежутки между нижними (как и нижние между верхними). Длинные клыки овальные в сечении, изогнуты назад и обладают мощными, глубоко сидящими корнями. Задний край клыка снабжен мелко зазубренным режущим гребнем, отходящим от вершины зуба к его основанию. На небе имеются особые углубления для вершин нижних клыков. В целом резцы и клыки очень мощные, и, очевидно, хорошо приспособленные к разрыванию даже крупной жертвы. Предклыковые зубы и клыки являлись в первую очередь хватательными, хотя гребни на краях верхних резцов, заходящие за края нижних, делали резцы отчасти и режущими. Наличие на предклыковых зубах зачаточных пяток указывает на их слабо выраженную измельчающую функцию (гораздо более совершенными в этом плане были резцы тапиноцефалоидей). Заклыковые зубы небольшие, слабо согнутые, наклоненные назад, с острыми и уплощенными с боков коронками, обладающими зазубренными режущими кантами. Редкие и сравнительно слабые, режущие по своему типу, они мало годились для измельчения пищи и, скорее всего, играли вспомогательную роль при удержании добычи. Поскольку жевать свою добычу титанофонеи не могли, они глотали ее целиком или отрывали куски мощным рывком головы. Ребра двухголовчатые. Лопатка, ключица и межключица узкие. Лопатка изогнута очень слабо; поскольку ее кривизна при близком расположении к грудной клетке в известной форме отражает ее форму, можно сделать вывод о сжатом с боков туловище (у крокодилов оно обычно уплощенное сверху вниз). Вентральная часть тазового пояса расширенная. В симфизе таза между лобковыми и седалищными костями имеется зона неполного окостенения. В вертлужной впадине между подвздошной, лобковой и седалищной костями окостенение неполное. Подвздошная кость по своему строению промежуточная между таковой древнейших рептилий, где она вытянута назад, и подвздошной костью более прогрессивных синапсид, у которых ее верхний отдел вытянут вперед, как у млекопитающих. Ранее с титанофонеями сближали и описанного по фрагментам черепа адметофонея (Admethophoneus karagalensis). На сегодняшний день его относят к примитивным горгонопсиям типа камагоргона (Kamagorgon ulanovi).
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Tags: Вымершие синапсиды, Пермь, терапсиды |
| 02:29 pm [industrialterro] [Link] |
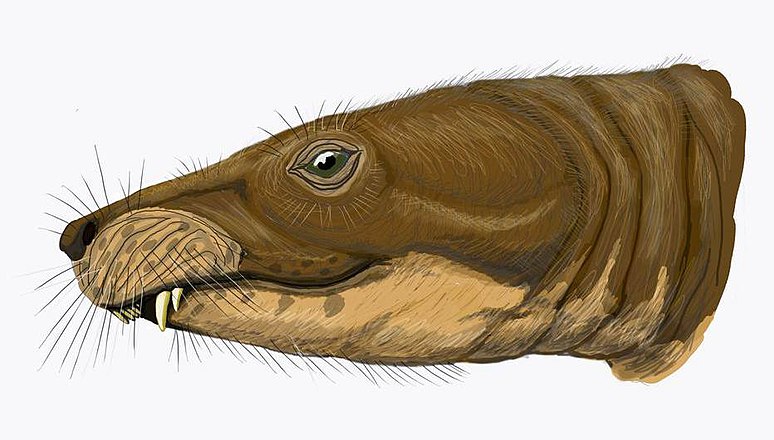
Theriognathus Териогнат (Theriognathus) – позднепермский тероцефал из семейства вайтсиид. Известен также под названиями Aneugomphius, Whaitsia, Notosollasia и др. В реальности все эти тероцефалы оказались представителями не только одного рода, но, вероятно, и одного вида - Theriognathus ictidoceps (или T. microps). Длина черепа от 8 до 31 см, общая длина до 2 метров. Морда относительно широкая, череп низкий. Хорошо развиты резцы, присутствуют 2 пары верхних клыков (вторая пара может быть замещающей). Верхние резцы нависают над нижними, направлены вниз и назад. Заклыковые зубы обычно отсутствуют. Твердого вторичного неба нет, но мягкое небо могло продолжаться далеко назад, помогая дышать во время еды. Предполагается, что вместо задних зубов могли быть роговые пластины, покрывавшие десны. Скелет массивный. Считается всеядным животным, но известна находка черепа (отнесенного к роду Notosollasia) с застрявшим между челюстей поврежденным черепом мелкого дицинодонта. Это позволяет предположить, что териогнаты были хищниками, которые при атаке хватали жертву за голову (так делает в наше время ягуар). Возможно, мелкая добыча проглатывалась целиком. Известен из позднепермских отложений (зоны Cistecephalus - Dicynodon) Южной Африки.
Tags: Вымершие синапсиды, Пермь, терапсиды |









































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}