[Recent Entries][Archive][Friends][User Info]
September 29th, 2011
| September 29th, 2011 | |
|---|---|
| 04:23 pm [industrialterro] [Link] |
Cheirolepis Cheirolepis ('hand fin') is an extinct genus of ray-finned fish that lived in the Devonian period of Europe and North America. It is the only genus yet known within the family Cheirolepidae and the order Cheirolepiformes. It was among the most basal of the Devonian actinopterygians and is considered the first to possess the "standard" dermal cranial bones seen in later actinopterygians. Cheirolepis was a predatory freshwater fish about 55 centimetres (22 in) long. It had a streamlined body with small, triangular ganoid scales similar to those of the Acanthodii. Cheirolepis had well-developed fins which gave it speed and stability, and was probably an active predator. Based on the size of its eyes, it hunted by sight. Cheirolepis's jaws, lined with sharp teeth, could be opened very wide, allowing it to swallow prey two thirds of its own size. Six possible species of Cheirolepis are currently known. The type species, named in 1835, is C. trailli. Remains of this species have been found from Scotland and date back to the Eifelian and Givetian stages of the Middle Devonian. C. canadensis was described in 1881 from material found in Miguasha, Canada that dated back to the middle Frasnian stage of the Late Devonian. Two species, C. gracilis and C. gaugeri, have been found from Germany and Belarus in deposits that are of Givetian age. They were first described in 1973 from scale material that is now of questionable validity. Another species has been found from Belarus that lived during the Eifelian, and has been named C. sinualis. A new species has recently been described from a locality in Red Hill, Nevada deposited during the Mid-Late Devonian boundary. The specimen from which this species was named, consisting of scales and a lower jaw, was originally referred to C. canadensis. New, more complete specimens have shown it to be distinct from the type, although a species name is yet to be given for the remains.
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие рыбы, Девон, Лучепёрые |
| Time | Event |
| 04:53 pm [industrialterro] [Link] |
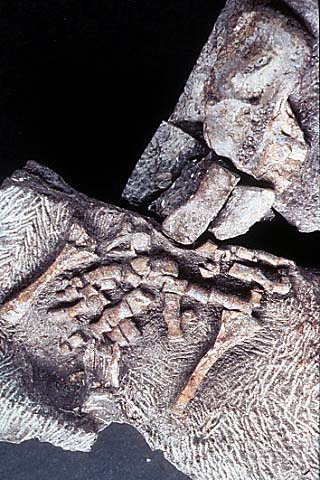
Acanthostega В девоне появляются первые амфибии. Акантостега (Acanthostega) — род ископаемых тетрапод, живших в позднем девоне и являющихся промежуточным звеном между лопастепёрыми рыбами и наземными животными. Одни из первых хордовых, развивших конечности. Окаменевший череп акантостеги (Acanthostega gunneri) был обнаружен в восточной Гренландии в 1933 году, однако описан лишь в 1952 году Эриком Ярвиком. В 1987 году Дженифер Кларк были обнаружены новые фрагменты нескольких особей. Кости акантостеги были найдены в древних речных отложениях, предполагается что именно в реках эти животные и обитали. Акантостега достигала длины около 60 см. Конечности не имели запястий, что говорит о крайне низкой приспособленности к передвижению по суше, и на каждой из них насчитывалось 8 пальцев. Строение скелета указывает на наличие внутренних жабр. Слабые конечности, которые не смогли бы выдержать вес животного, и короткие рёбра, на которые оно также не могло опираться, говорят о его преимущественно водном образе жизни. Acanthostega had eight digits on each hand (the number of digits on the feet is unclear) linked by webbing, it lacked wrists, and was generally poorly adapted to come onto land. Acanthostega also had a remarkably fish-like shoulder and forelimb. The front foot of Acanthostega could not bend forward at the elbow, and thus could not be brought into a weight bearing position, appearing to be more suitable for paddling or for holding on to aquatic plants. It had lungs, but its ribs were too short to give support to its chest cavity out of water, and it also had gills which were internal and covered like those of fish, not external and naked like those of some modern amphibians which are almost wholly aquatic. Acanthostega is the first tetrapod to show the shift in locomotory dominance from the pectoral to pelvic girdle. There are many morphological changes that allowed the pelvic girdle of Acanthostega to become a weight-bearing structure. In more ancestral states the two sides of the girdle were not attached. In Acanthostega there in contact between the two dies and fusion of the girdle with the sacral rib of the vertebral column. These fusions would have made the pelvic region more powerful and equipped to counter the force of gravity when not supported by the buoyancy of and aquatic environment. Research based on analysis of the suture morphology in its skull indicates that the species may have bitten directly on prey at or near the water's edge. Markey and Marshall compared the skull with the skulls of fish, which use suction feeding as the primary method of prey capture, and creatures known to have used the direct biting on prey typical of terrestrial animals. Their results indicate that Acanthostega was adapted for what they call terrestrial-style feeding, strongly supporting the hypothesis that the terrestrial mode of feeding first emerged in aquatic animals. If correct, this shows an animal specialized for hunting and living in shallow waters in the line between land and water. Newer research also indicates that it is possible Acanthostega actually evolved from an ancestro that was more terrestrail adapted than itself. Acanthostega is seen as part of widespread speciation in the late Devonian period, starting with purely aquatic lobe-finned fish, with their successors showing increased air-breathing capability and related adaptions to the jaws and gills, as well as more muscular neck allowing freer movement of the head than fish have, and use of the fins to raise the body of the fish. These features are displayed by the earlier Tiktaalik, which like the Ichthyostega living around the same time as Acanthostega showed signs of greater abilities to move around on land, but is thought to have been primarily aquatic.
Репродукции (1, 2, 3, 4, 5, 6):
Ископаемые останки (1, 2, 3, 4, 5, 6):
Tags: Вымершие амфибии, Девон, Ихтиостегалии, Лабиринтодонты |
| Time | Event |
| 05:15 pm [industrialterro] [Link] |
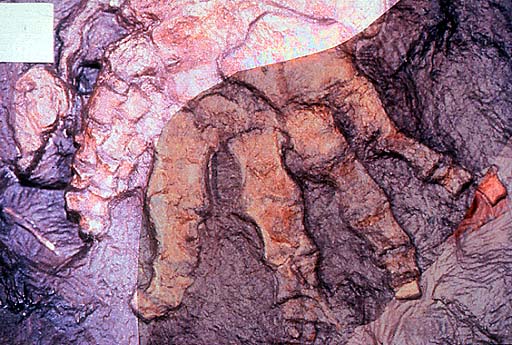
Ichthyostega Ихтиосте́га (лат. Ichthyostega) — род ранних тетрапод, живший в верхнем девонском периоде, около 367—362,5 млн лет назад, и представляющий собой первое промежуточное звено между рыбами и амфибиями. Этот род рассматривается в составе амфибий, однако он не является прямым предком современных видов, предки которых — лепоспондилы (Lepospondyli) — появились в каменноугольном периоде. У ихтиостегий были ноги, но их конечности, возможно, не использовались для ходьбы, а со временем были использованы для преодоления болот. Ихтиостеги имели хвостовой плавник и некоторые органы чувств, функционирующие только в воде. Тело их было покрыто мелкими чешуйками. По мнению некоторых учёных ихтиостеги наряду с определёнными видами кистепёрых рыб определили направление филогенеза в сторону возникновения наземных позвоночных. В 1932 году Сейв-Содерберг описал четыре вида ихтиостегий из верхнего девона, найденных в восточной части Гренландии и один вид принадлежащий роду Ichthyostegopsis, I. wimani. Эти виды могли быть синонимичными, потому что их морфлогические различия не были резко выражены. Виды различались в пропорциях и строении черепа. Сравнение было проведено на четырнадцати экземплярах, собранных в 1931 году Датской Восточной Гренландской Экспедицией. Дополнительные экземпляры были собраны между 1933 и 1955 годами. Род Ихтиостега находится в близком родстве с акантостегой (Acanthostega gunnari), также обнаруженной в восточной Гренландии. По сравнению с акантостегой череп ихтиостеги выглядит более рыбообразным, однако её пояс передних конечностей сильнее и лучше адаптирован к наземной жизни. Ихтиостеги были около 1,5 метров длиной и имели по семь пальцев на задних ногах. Точное количество пальцев на передних лапах пока не установлено, но вероятно, что их было тоже семь. На хвосте у них был плавник «рыбьего» типа поддерживаемый невральными и гемальными дугами. Ноздри располагались у нижнего края челюстей. Слезная кость примыкает к ноздре, но не к глазнице. Межвисочная кость отсутствует (это обстоятельство исключает возможность происхождения большинства темноспондилов от этого рода). Заднетеменная кость непарная. Челюстная кость соприкосается с квадратноскуловой. Сохраняются подкрышечные и предкрышечные кости. Носовые кости широкие. Глазницы овальные и располагаются в центральной части черепа. Хорда через отико-окципитальную часть мозговой коробки доходит до гипофизарной ямы. Early tetrapods like Ichthyostega and Acanthostega differed from animals like Crossopterygians (for instance Eusthenopteron or Panderichthys) in their increased adaptations for life on land. Though Crossopterygians possessed lungs, they used gills as their primary means of acquiring oxygen; Ichthyostega appears to have relied on its lungs as its primary apparatus for breathing. The skin of early tetrapods, unlike that of Crossopterygians, helped retain bodily fluids and deter desiccation. Crossopterygians used their body and tail for locomotion and their fins for balance; Ichthyostega used its limbs for locomotion and its tail for balance. The size of an adult Ichthyostega (1.5 m) precluded completely terrestrial locomotion. Yet the massive ribcage was made up of overlapping ribs and the animal possessed a stronger skeletal structure, a more rigid spine, and forelimbs apparently powerful enough to pull the body from the water. These anatomical modifications clearly evolved to handle the lack of buoyancy experienced on land. The hindlimbs were smaller than the forelimbs and unlikely to have born full weight in an adult, while the broad, overlapping ribs would have inhibited side-to-side movements. Jennifer A. Clack suggests that Ichthyostega and its relatives spent time basking in the sun to raise their body temperatures, much as some animals do today: the Marine Iguanas on the Galapagos Island or the Gharial. They would have returned to the water to cool themselves, hunt for food and reproduce. A lifestyle that required strong forelimbs to pull at least their anterior part out of the water, and a stronger ribcage and spine to support them while sunbathing on their abdomen like modern crocodiles. New studies suggests that the juveniles were more aquatic than the adults, and the possibility that Ichthyostega came out of the water only as a fully mature adult. Water was also still a requirement, because the gel-like eggs of the earliest terrestrial tetrapods couldn't survive out of water, so reproduction could not occur without it. Water was also needed for their larvae and external fertilization. Most land-dwelling vertebrates have since developed two methods of internal fertilization; either direct as seen in all amniotes and a few amphibians, or indirect for many salamanders by placing a spermatophore on the ground which then is picked up by the female salamander. The Ichthyostegalians (Elginerpeton, Acanthostega, Ichthyostega, etc.) were succeeded by temnospondyls and anthracosaurs, such as Eryops, amphibians that truly developed the ability to walk on land. Until 2002, there was a gap of 20 million years between the two groups ( Romer's Gap). In 2002 a 350 million year old fossil from the lower Mississippian, Pederpes finneyae was described and helped to close the gap: it is the earliest-known tetrapod to show the beginnings of terrestrial locomotion.
Репродукции (1, 2, 3, 4, 5, 6):
(Кстати, плаун, что показан тут на среднем плане был высотой до 8 метров.)
Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие амфибии, Девон, Ихтиостегалии, Лабиринтодонты |
| Time | Event |
| 05:37 pm [industrialterro] [Link] |
Elginerpeton Elginerpeton is a monotypic genus of early tetrapod, the fossils of which were recovered from Scat Craig, Scotland, in rocks dating to the late Devonian Period (Frasnian stage, 395 million years ago). The only known fossil has been given the name Elginerpeton pancheni. Elginerpeton is known from a partial skeleton including a partial shoulder and hip, a femur, tibia (lower hind limb), and jaw fragments. It is estimated to have measured about 1.5 m (5 ft) in length.
Tags: Вымершие амфибии, Девон, Ихтиостегалии, Лабиринтодонты |
| Time | Event |
| 05:57 pm [industrialterro] [Link] |
Hynerpeton Hynerpeton (pronounced /haɪˈnɜrpətɒn/, from Greek Υνηρπετον "creeping animal from Hyner") was a basal carnivorous tetrapod that lived in the lakes and estuaries of the Late Devonian period around 360 million years ago. Like many primitive tetrapods, it is sometimes referred to as an "amphibian", though it is not a true member of the modern Lissamphibia. The Late Devonian saw the evolution of plants into trees and growing into vast forests pumping oxygen into the air, possibly giving Hynerpeton an edge because it evolved complex lungs to exploit it. Its lungs probably consisted of sacs like modern terrestrial vertebrates. In 1993, the paleontologists Ted Daeschler and Neil Shubin found the first Hynerpeton fossil, a shoulder bone, near Hyner, Pennsylvania. They were surveying the Devonian rocks of Pennsylvania in search of fossil evidence for the origin of animal limbs. The animal had a very robust shoulder, which indicated that it had powerful appendages. Only a few bones have been found from Hynerpeton, in Red Hill, Pennsylvania, U.S.A.. The known fossils include two shoulder girdles, two lower jaws, a jugal bone and some gastralia. The structure of the shoulder girdle indicates this animal may have been one of the earlier, more primitive tetrapods to evolve during the Devonian. Information on the relationship of the known fossils of Hynerpeton to other Devonian tetrapods can be found in Gaining Ground The Origin and Evolution of Tetrapods by Jennifer A. Clack. It is thought that that these early amphibians are descended from lobe-finned fish, such as Hyneria, whose stout fins evolved into legs and their swim bladder into lungs. It is still not known whether Hynerpeton is the direct ancestor to all later backboned land animals (including humans), but the fact that it had eight fingers, not five, suggests that it is simply our evolutionary cousin.
Tags: Вымершие амфибии, Девон, Ихтиостегалии, Лабиринтодонты |
| Time | Event |
| 06:11 pm [industrialterro] [Link] |
Tulerpeton Тулерпетон (Tulerpeton) — ископаемый род позднедевонских амфибий. Был впервые обнаружен в Тульской области, от которой и получил своё имя. Известен по практически польностью сохранившимся плечевому поясу, передним и задним конечностям, фрагментам черепа и другим мелким находкам. Тулерпетон достигал 50 см в длину, имел по 6 пальцев на каждой конечности и полностью утратил внутренние жабры. Строение скелета говорит о том, что он был более приспособленным к жизни на суше чем Акантостега, однако его ноги больше подходили для проталкивания туловища по мягкому влажному грунту, чем для ползания по земле. Находки тулерпетона были сделаны в морских отложениях, в то время как остальные ранние амфибии найдены в пресноводных. Это, возможно, указывает на длительное сохранение у амфибий физиологических механизмов адаптации к жизни в солёной воде и возможности пересекать моря, что позволило им распространиться уже в девоне вдоль всего экваториального пояса Земли. Tulerpeton lived approximately 365 million years ago, in the Late Devonian period when the climate was fairly warm and there were no glaciers. Land had already been colonized by plants. During the Devonian period, terrestrial tetrapods – the ancestors of present day reptiles, birds, amphibians and mammals - first began to appear. Even though Tulerpeton breathed air, it lived mainly in shallow marine water. The Andreyevka fossil bed where it was discovered was at least 200 km from the nearest landmass during this era. The fossils of plants in the area tell us that the salinity of the waters where it lived fluctuated wildly, indicating that the waters were quite shallow. Because the bones of the neck and the pectoral girdle were disconnected, Tulerpeton could lift its head. Therefore, in shallow water, it had a considerable advantage over the other animals whose heads only moved side to side. The later land animals that descended from Tulerpeton’s relatives needed this head flexion on land, but the condition probably evolved because of the advantage that this gave it in shallow marine waters, not for land. In the book “Vertebrate Life”, authors Pough, Janis, and Heiser say that,” The development of a distinct neck, with the loss of the opercular bones and the later gain of a specialized articulation between the skull and the vertebral column (not yet present in the earliest tetrapods), may be related to lifting the snout out of the water to breath air or to snap at prey items.” The six fingered hands and toes were stronger than the fins from which they developed, therefore “tulerpeton” had an advantage in propelling itself through shallow and brackish water, but the limbs do not yet seem strong enough for extensive use on land.
Tags: Антракозавры, Вымершие амфибии, Девон, Лабиринтодонты |
| Time | Event |
| 07:48 pm [industrialterro] [Link] |
Chitinozoa Chitinozoa (singular: chitinozoan, plural: chitinozoans) are a taxon of flask-shaped, organic walled marine microfossils produced by an as yet unknown animal. Common from the Ordovician to Devonian periods (i.e. the mid-Paleozoic), the millimetre-scale organisms are abundant in almost all types of marine sediment across the globe. This wide distribution, and their rapid pace of evolution, makes them valuable biostratigraphic markers. Their bizarre form has made classification and ecological reconstruction difficult. Since their discovery in 1931, suggestions of protist, plant, and fungal affinities have all been entertained. The organisms have been better understood as improvements in microscopy facilitated the study of their fine structure, and there is mounting evidence to suggest that they represent either the eggs or juvenile stage of a marine animal. The ecology of chitinozoa is also open to speculation; some may have floated in the water column, where others may have attached themselves to other organisms. Most species were particular about their living conditions, and tend to be most common in specific paleoenvironments. Their abundance also varied with the seasons. Chitinozoa range in length from around 50 to 2000 micrometres. They appear dark to almost opaque when viewed under an optical microscope. External ornamentation is often preserved on the surface of the fossils, in the form of hairs, loops or protrusions, which are sometimes as large as the chamber itself. The range and complexity of ornament increased with time, against a backdrop of decreasing organism size. The earliest Ordovician species were large and smooth-walled; by the mid-Ordovician a large and expanding variety of ornament, and of hollow appendages, was evident. While shorter appendages are generally solid, larger protrusions tend to be hollow, with some of the largest displaying a spongy internal structure. However, even hollow appendages leave no mark on the inner wall of the organisms: this may suggest that they were secreted or attached from the outside. There is some debate about the number of layers present in the organisms' walls: up to three layers have been reported, with the internal wall often ornamented; some specimens only appear to display one. The multitude of walls may indeed reflect the construction of the organism, but could be a result of the preservational process. Immature" or juvenile examples of Chitinozoans have not been found; this may suggest that they didn't "grow", that they were moults (unlikely), or that the fossilisable parts of the organism only formed after the developmental process was complete. Most chitinozoans are found as isolated fossils, but chains of multiple tests, joined from aperture to base, have been reported from all genera. Very long chains tend to take the form of a spring. Occasionally, clusters or condensed chains are found, packed in an organic "cocoon". It is not immediately clear what mode of life was occupied by these improbably shaped fossils, and an answer only becomes apparent after following several lines of reasoning. The fossils' restriction to marine sediments can be taken as sound evidence that the organisms dwelt in the Palæozoic seas - which presents three main modes of life:
An infaunal mode of life can be quickly ruled out, as the fossils are sometimes found in alignment with the depositing current; as nothing attached them to the bottom, they must have fallen from the water column. The ornament of the chitinozoans may cast light on the question. Whilst in some cases a defensive role - by making the vessel larger, and thus less digestible by would-be predators - seems probable, it is not impossible that the protrusions may have anchored the organisms to the sea floor. However, their low-density construction makes this unlikely: perhaps more plausible is that they acted to attach to other organisms. Longer spines also make the organisms more buoyant, by decreasing their Rayleigh number (i.e. increasing the relative importance of water's viscosity) — it is therefore possible that at least the long-spined chitinozoans were planktonic "floaters". On the other hand, the walls of some chitinozoans were probably too thick and dense to allow them to float. Whilst little is known about their interactions with other organisms, small holes in the tests of some chitinozoans are evidence that they were hosts to some parasites.[5][9][10] Although some forms have been reinterpreted as "pock-marks" caused by the disintegration of the diagenetic mineral pyrite, the clustering of cylindrical holes around the chamber — where the flesh of the organism was likely to be concentrated — is evidence for a biological cause. Corals in Gotland with daily growth markings have been found in association with abundant chitinozoans, which allow the detection of seasonal variation in chitinozoan abundance. A peak in abundance during the late autumn months is observed, with the maxima for different species occurring on different dates. Such a pattern is also observed in modern-day tropical zooplankton. The diversity of living habits is also reflected by the depth of water and distance from the shore. Different species are found in highest abundance at different depths. While deeper waters around 40 km from the shoreline are generally the optimal environment, some species appear to prefer very shallow water. On the whole, chitinozoans are less abundant in turbulent waters or reef environments, implying an aversion to such regimes when alive, if it is not an effect of sedimentary focusing. Chitinozoans also become rarer in shallower water - although the reverse is not necessarily true. They cannot survive freshwater input.
Tags: Вымершие одноклеточные, Девон |
| Time | Event |
| 08:04 pm [industrialterro] [Link] |
Немного девонской ботвы Эволюция наземной растительности девонского периода уже вполне определенно выявлена. Отложения нижнего и среднего девона характеризуются еще низкоорганизованной растительностью, известной под названием «псилофитовой», наиболее типичным представителем которой является примитивное папоротиикообразное — псилофитон.
Tags: Девон, Растительность |
| Time | Event |
| 08:13 pm [industrialterro] [Link] |
Девонское вымирание Девонское вымирание — вымирание видов в позднем Девоне, было одним из крупнейших вымираний в истории земной флоры и фауны. Основное вымирание произошло на границе, которая отмечает начало последней фазы Девонского периода, Фаменского (Famennian) экологического века, (Фраснско — Фаменская (Frasnian-Famennian) граница), около 374 миллионов лет назад, когда почти все бесчелюстные рыбы неожиданно исчезли (их окаменелости в отложениях перестают встречаться). Второй сильный опустошающий импульс завершил Девонский период. Повсюду вымерло 19 % семейств и 50 % всего генофонда. Хотя понятно, что имело место массовое сокращение биоразнообразия к концу Девона, интервал времени, в течение которого это событие происходило, неясен: оценки варьируются от 500 тысяч до 15 миллионов лет, в последнем случае оно продолжалось в течение всего Фаменского века (Famennian). Не совсем понятно, было ли это событие представлено двумя резкими скачками (пиками) массового вымирания или серией из меньших по размеру вымираний, однако результаты последнего исследования скорее указывают на многоэтапное развитие вымирания, из серии отдельных импульсов вымирания на протяжении временного интервала около трёх миллионов лет. Некоторые предполагают, что вымирание состояло по меньшей мере из семи отдельных событий, происходивших в течение 25 миллионов лет, включая наиболее заметные вымирания в конце Живетского (Givetian), Фраснского (Frasnian) и Фаменского (Famennian) веков. Некоторые ссылаются на 250-миллионолетний диапазон, в течение которого вымирания имели место. К позднему Девону суша была полностью освоена и заселена растениями, насекомыми и земноводными, а моря и океаны были полны рыбой. Кроме того, в этот период уже существовали гигантские рифы, сформированные кораллами и строматопороидами. Евроамериканский континент и Гондвана только начали движение друг к другу, чтобы в будущем сформировать суперконтинент Пангею. Вероятно, вымирание в основном затронуло морскую жизнь. Рифообразующие организмы были почти полностью уничтожены, в итоге коралловые рифы возродились только с развитием современных кораллов в Мезозое. Брахиоподы (плеченогие), трилобиты и другие семейства тоже были тяжело затронуты. Причины этого вымирания пока неясны. Основная теория предполагает, что изменения в уровне океана и обеднение океанических вод кислородом послужили главной причиной вымирания жизни в океанах. Возможно, что активатором этих событий послужило глобальное похолодание или обширный океанический вулканизм, хотя падение внеземного тела, такого как комета, тоже вполне возможно. Некоторые статистические исследования морской фауны того времени наводят на мысль, что уменьшение разнообразия было связано, скорее, со спадом в темпе видообразования, чем ростом вымирания.
Tags: Девон, Массовое вымирание |
| Time | Event |
| 09:09 pm [industrialterro] [Link] |
Каменноугольный период Каменноу́гольный пери́од, сокращенно карбо́н (С) — геологический период в верхнем палеозое 360—299 млн лет назад. Назван из-за эпохи углеобразования в это время. Впервые появляются очертания величайшего суперконтинента в истории Земли — Пангеи. Пангея образовалась при столкновении Лавразии (Северная Америка и Европа) с древним южным суперконтинентом Гондваной. Незадолго до столкновения Гондвана повернулась по часовой стрелке, так что ее восточная часть (Индия, Австралия, Антарктида) переместилась к югу, а западная (Южная Америка и Африка) оказалась на севере. В результате поворота на востоке появился новый океан — Тетис, а на западе закрылся старый — океан Рея. В то же время океан между Балтикой и Сибирью становился все меньше; вскоре эти континенты тоже столкнулись. В отложениях этого периода находят огромные залежи каменного угля. Отсюда и произошло название периода. Есть и другое его наименование— карбон. Каменноугольный период подразделяется на три отдела: нижний, средний и верхний. В этот период физико-географические условия Земли претерпели значительные изменения, Неоднократно менялись очертания материков и морей, возникали новые горные хребты, моря, острова. В начале карбона происходит значительное опускание суши. Морем были затоплены обширные площади Атлантии, Азии, Рондваны. Уменьшилась площадь больших островов. Исчезли под водой пустыни северного континента. Климат стал очень теплым и влажным, В нижнем карбоне начинается интенсивный горообразовательный процесс: формируются Арденны, Гари, Рудные горы, Судеты, Атласскпе горы, Австралийские Кордильеры, Западно-Сибирские горы. Море отступает. В среднем карбоне суша вновь опускается, но гораздо меньше, чем в нижнем. В межгорных котловинах накапливаются мощные толщи континентальных отложений. Формируется Восточный Урал, Пеннинскис горы. В верхнем карбоне море вновь отступает. Значительно уменьшаются внутриконтинентальные моря. На территории Гондваны возникают большие ледники, в Африке и Австралии—несколько меньшие. В конце карбона в Европе и Северной Америке климат претерпевает изменения, становится частично умеренным, а частично жарким и сухим. В это время происходит формирование Центрального Урала. Усиленная вулканическая деятельность в карбоне привела к насыщению атмосферы углекислым газом. Вулканический пепел, являющийся чудесным удобрением, сделал плодородными карбоновые почвы. Теплый и влажный климат господствовал на материках длительное время. Все это создало чрезвычайно благоприятные условия для развития наземной флоры, в том числе высших растений каменноугольного периода—кустов, деревьев и травянистых растений, жизнь которых была тесно связана с водой. Они росли главным образом среди громадных болот и озер, близ солоновато водных лагун, на побережье морей, на влажной илистой почве. По образу жизни они походили на современных мангровых, которые растут на низинных берегах тропических морей, в устьях больших рек, в болотистых лагунах, поднимаясь над водой на высоких корнях-ходулях. Значительное развитие в каменноугольный период получили плауновидные, членистостебельные и папоротниковые, давшие большое количество древовидных форм. Древовидные плаунообразные достигали 2 м в диаметре и 40 м в высоту. У них еще не было годичных колец. Пустой ствол с мощной разветвленной кроной надежно удерживался в рыхлой почве большим корневищем, разветвлявшимся на четыре главные ветви. Эти ветви в свою очередь дихотомически разделялись на корневые отростки. Их листья, до метра в длину, густыми султанообразными пучками украшали концы ветвей. На концах листьев располагались почки, в которых развивались споры. Стволы плаунообразных были покрыты чешуей—рубцами. К ним прикреплялись листья. В этот период были распространены гигантские плаунообразные—лепидодендроны с ромбическими рубцами на стволах и сигиллярии с шестигранными рубцами. В отличие от большинства плаунообраз-.ныху сигиллярий был почти неразветвленный ствол, на котором вырастали спорангии. Среди плаунообразных были и травянистые растения, полностью вымершие в пермский период. Членистостебельные растения разделяются на две группы: клинолистые и каламиты. Клинолистые являлись водными растениями. У них был длинный, членистый, слегка ребристый стебель, к узлам которого кольцами прикреплялись листья- Почковидные образования содержали споры. На воде клинолистые держались с помощью длинных разветвленных стеблей, похожих на современный водяной лютик. Клинолистые появились в среднем девоне и вымерли в пермский период. Каламиты были древовидными растениями до 30 м высотой. Они образовывали болотные леса. Некоторые виды каламитов проникли далеко на материк. Их древние формы имели дихотомические листья. Впоследствии преобладали формы с простыми листьями и годичными кольцами. Эти растения имели сильно разветвленное корневище. Нередко из ствола вырастала дополнительные корни и ветви, покрытые листьями. В конце карбона появляются первые представители хвощевых—небольшие травянистые растения. Среди карбоновой флоры видную роль играли папоротники, в частности травянистые, но своему строению напоминавшие псилофитов, и настоящие папоротники—большие древовидные растения, корневищем закреплявшиеся в мягком грунте. У них был шершавый ствол с многочисленными ветвями, на которых росли широкие папоротниковидные листья. Голосеменные карбоновых лесов относятся к подклассам семенных папоротников и стахиоспермид. Их плоды развивались на листьях, что является признаком примитивной организации. В то же время у линейных или ланцетных листьев голосеменных было довольно сложное жилкоианне. Наиболее совершенные растения карбона — кордаиты. Их цилиндрические безлистые стволы до 40 м , высотой разветвлялись. Ветви имели на концах широ-, кие линейные или ланцетные листья с сетчатым жилкованием|. Мужские спорангии (микроспорангии) имели вид почек. Из женских спорангиев развивались ореховидные плоды. Результаты микроскопического исследования плодов показывают, что эти растения, похожие на цикадовых, являлись переходными формами к хвойным растениям. В каменноугольных лесах появляются первые грибы, моховидные растения (наземные и пресноводные), образовывавшие иногда колонии, н лишайники. Распространение одних и тех же каменноугольных растений от Северной Америки до Шпицбергена свидетельствует о том, что от тропиков до полюсов господствовал относительно равномерный теплый климат, на смену которому в верхнем карбоне пришел довольно прохладный. В прохладном климате росли голосеменные папоротники и кордаиты. Отмирая, растения каменноугольного периода падали в воду, их заносило илом, и, пролежав миллионы лет, они постепенно превратились в уголь. Каменный уголь образовывался изо всех частей растения: древесины, коры, ветвей, листьев, плодов. В уголь превращались и остатки животных. Об этом свидетельствует тот факт, что остатки пресноводных и наземных животных в карбоновых отложениях встречаются относительно редко. Морской животный мир карбона характеризовался разнообразием видов. Чрезвычайно распространенными были фораминиферы, в частности фузулиниды с веретеновидными раковинами величиной с зерно. В среднем карбоне появляются швагерины. Их шаровидная раковина была величиной с небольшую горошину. Из раковин фораминифер позднего карбона в отдельных местах образовались залежи известняков. Среди кораллов еще встречались немногочисленные роды табулят, но начинали преобладать хететиды. Одиночные кораллы нередко имели толстые известковые стенки, Колониальные кораллы образовывали рифы. В это время интенсивно развиваются иглокожие, в частности морские лилии и морские ежи. Многочисленные колонии мшанок иногда образовывали мощные известняковые отложения. Чрезвычайно развились плеченогие моллюски, в частности продуктусы, по приспособляемости и географическому распространению намного превосходившие всех встречающихся на Земле плеченогих. Величина их раковин достигала 30 см в поперечнике. Одна створка раковины была выпуклой, а другая—в виде плоской крышечки. Прямой удлиненный замочный край часто имел полые шипы. У.некоторых форм продуктусов шипы в четыре раза превышали диаметр раковины. С помощью шипов продуктусы держались на листьях водных растений, переносивших их по течению. Иногда своими шипами они прикреплялись к морским лилиям или водорослям и жили возле них в висячем положении. У рихтофений одна створка раковины преобразовалась в рог длиной до 8 см. В каменноугольный период почти полностью вымирают наутилоидеи, за исключением наутилусов. Данный род, распавшийся на 5 групп (которые были представлены 84 видами), дожил до нашего времени. Продолжают существовать ортоцерасы, раковины которых имели резко выраженное внешнее строение. Роговидно согнутые раковины циртоцерасов почти не отличались от раковин их девонских предков. Аммониты были представлены двумя отрядами— гониатитами и агониатитами, как и в девонском периоде двустворчатые моллюски — одномышечными формами. Среди них — множество пресноводных форм, населявших карбоновые озера и болота. Появляются первые наземные гастроподы —животные,дышавшие легкими. Трилобиты достигли значительного расцвета в ордовикский и силурийский периоды. В каменноугольный период сохранились лишь немногочисленные их роды и виды. К концу каменноугольного периода трилобиты почти полностью вымерли. Этому способствовало то, что головоногие моллюски и рыбы питались трилобитами и потребляли ту же пищу, что и трилобиты. Строение тела трилобитов было несовершенным: панцирь не защищал брюха, конечности были небольшими и слабыми. Трилобиты не имели органов нападения. Некоторое время они могли защищаться от хищников, свертываясь наподобие современных ежей. Но в конце карбона появились рыбы с мощными челюстями, разгрызавшими их панцирь. Поэтому от многочисленного типа инерми сохранился только один род. В озерах каменноугольного периода появляются ракообразные, скорпионы, насекомые. Карбоновые насекомые обладали признаками многих родов современных насекомых, поэтому отнести их к какому .либо одному известному нам теперь роду невозможно. Несомненно, предками насекомых каменноугольного периода были ордовикские трилобиты. У девонских и силурийских насекомых было много общего с некоторыми их предками. Они играли уже значительную роль в животном мире. Однако подлинного расцвета насекомые достигли в каменноугольном периоде. Представители самых малых из известных видов насекомых были 3 см в длину; размах крыльев у самых крупных (например, у стенодиктии) достигал 70 см, у древней стрекозы меганевры—одного метра. Тело меганевры имело 21 сегмент. Из них 6 составляли голову, 3—грудь с четырьмя крыльями, 11—брюшко, конечный сегмент походил на шиловидное продолжение хвостового щита трилобитов. Многочисленные пары конечностей были расчленены. С их помощью животное и ходило, и плавало. Молодые меганевры обитали в воде, превращаясь во взрослых насекомых в результате линьки. У меганевры были крепкие челюсти и сложные глаза. В верхнем каменноугольном периоде древние насекомые вымерли, их потомки были более приспособленными к новым условиям жизни. Прямокрылые в ходе эволюции дали термитов и стрекоз, эвриптерусы—муравьев. Большинство древних форм насекомых переходили к наземному образу жизни лишь в зрелом возрасте. Размножались они исключительно в воде. Таким образом, перемена влажного климата на более сухой для многих древних насекомых была катастрофой. В каменноугольном периоде появляется много акул. Это были еще не настоящие акулы, населяющие современные океаны, однако по сравнению с другими группами рыб, они представляли собой самых совершенных хищников. В отдельных случаях их зубы и плавниковые типы переполняют каменноугольные отложения. Это свидетельствует, о том, что каменноугольные акулы жили в любой воде. Зубы зазубренные, широкие, режущие, бугристые, так как акулы питались разнообразнейшими животными. Постепенно они истребили примитивных девонских рыб. Ножевидные зубы акул легко разгрызали панцирь трилобитов, широкие бугристые зубные пластинки хорошо раздробляли толстые раковины моллюсков. Пиловидиые, заостренные ряды зубов позволяли акулам питаться колониальными животными. Формы и размеры акул были такими же разнообразными, как и способ их питания. Одни из них окружали коралловые рифы и с молниеносной быстротой преследовали свою добычу, другие же неторопливо охотились на моллюсков, трилобитов либо зарывались в ил и подстерегали добычу. Акулы с пиловидным выростом на голове искали жертв в зарослях морских водорослей. Крупные акулы часто нападали на более мелких, поэтому у некоторых из последних для защиты в ходе эволюции возникли плавниковые шипы и накожные зубы. Акулы интенсивно размножались. Это в конечном итоге привело к перенаселению моря этими животными. Были истреблены многие формы аммонитов, исчезли одиночные кораллы, представлявшие для акул легкодоступную питательную пищу, значительно сократилось число трилобитов, погибли все моллюски, имевшие тонкую раковину. Лишь толстые раковины спириферов не поддавались хищникам. Сохранились и продуктусы. Они защищались от хищников длинными шипами. В пресноводных бассейнах каменноугольного периода жило множество эмалевочешуйчатых рыб. Некоторые из них прыгали по илистому берегу, наподобие современных рыб-прыгунов. Спасаясь от врагов, насекомые покинули водную среду и заселили сушу сначала вблизи болот и Озер, а затем—горы, долины и пустыни каменноугольных материков. Среди насекомых каменноугольного периода отсутствуют пчелы и бабочки. Это понятно, так как в то время еще не было цветковых растений, чьей пыльцой и нектаром питаются эти насекомые. Животные, дышащие легкими, впервые появляются на материках девонского периода. Ими были земноводные. Жизнь земноводных тесно связана с водой, поскольку размножаются они лишь в воде. Теплый влажный климат карбона чрезвычайно благоприятствовал расцвету земноводных. Их скелеты еще не полностью окостенели, челюсти имели нежные зубчики. Кожа была покрыта чешуей. За низкий крышеобразный череп вся группа земноводных получила наименование стегоцефалов (панцироголовых). Размеры тела земноводных колебались от 10 см до 5м. У большинства из них было четыре ноги с короткими пальцами, Некоторые имели когти, позволявшие им лазить по деревьям. Появляются и безногие формы. В зависимости от образа жизни земноводные приобрели тритоновидную, змеевидную, саламандровидную формы. В черепе земноводных было пять отверстий: два носовых, два глазных и теменной глаз. Впоследствии этот теменной глаз преобразовался в шишковидную железу головного мозга млекопитающих. Спина у стегоцефалов была голой, а брюхо покрывала нежная чешуя. Они населяли неглубокие озера и болотистые места близ побережья. Наиболее характерный представитель первых пресмыкающихся—эдафозавр. Он напоминал огромную ящерицу. На спине у него имелся высокий гребень из длинных костяных шипов, соединенных между собой кожистой перепонкой. Эдафозавр был травоядным ящером и жил вблизи каменноугольных болот. С каменноугольными отложениями связано большое количество каменноугольных бассейнов, залежи нефти, железа, марганца, меди, известняков. Длился этот период 65 млн. лет.
Репродукции (1, 2, 3, 4, 5, 6, 7):
Tags: Карбон |


















































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Previous Day | 2011/09/29 [Archive] |
Next Day |