[Recent Entries][Archive][Friends][User Info]
April 24th, 2015
| April 24th, 2015 | |
|---|---|
| 01:26 pm [industrialterro] [Link] |
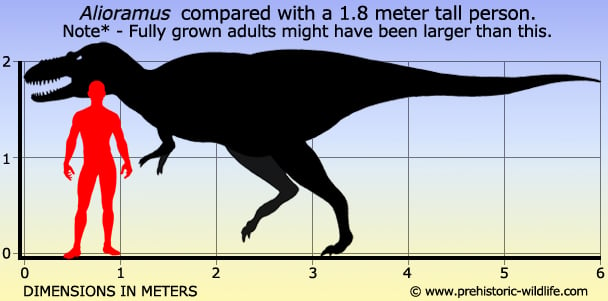
Alioramus Алиорам (Alioramus, от лат. alius ramus «другая ветвь») — род хищных динозавров из семейства тираннозавридов, живших в меловом периоде (около 70—65 млн лет назад) на территории современной пустыни Гоби (Монголия). Известны два вида. Впервые алиорама (A. remotus) описал советский палеонтолог С. М. Курзанов в 1976 году, обнаружив в монгольских отложениях фрагменты черепа и три плюсневые кости, принадлежащие динозавру, который жил 70-65 млн лет назад. Название от лат. alius ramus «другая ветвь» было выбрано из-за анатомических отличий алиорама от других тираннозаврид. С тех пор вопрос о принадлежности алиорама к тираннозавридам оставался дискуссионным. Новые останки обнаружены в монгольской пустыне Гоби в 2001 году сотрудниками Американского музея естественной истории под руководством Марка Норелла и Майкла Новачека. Они, вероятно, принадлежат девятилетнему подростку, который вырос всего на 85 %, жил 85—65 млн лет назад, достигал в длину 5 метров и весил около 369 кг. В 2009 году данные останки окончательно отнесли к новому виду — Alioramus altai. Главное отличие алтайского алиорама — необычные рога. Помимо обычных для тираннозавридов надбровных коротких рогов у него имеется по два пятисантиметровых рога, торчащих из каждой щеки. Использовать их для борьбы невозможно, поэтому предполагается, что рога служили украшением для привлечения самок. Алиорам по сравнению тарбозавром обладал маленьким черепом, миниатюрными зубками и слабыми челюстными мышцами и занимал отличную от последнего экологическую нишу. Alioramus (/ˌæli.ɵˈreɪməs/; meaning 'different branch') is a genus of tyrannosaurid theropod dinosaurs from the Late Cretaceous period of Asia. The type species, A. remotus, is known from a partial skull and three foot bones recovered from Mongolian sediments which were deposited in a humid floodplain about 70 million years ago. These remains were named and described by Soviet paleontologist Sergei Kurzanov in 1976. A second species, A. altai, known from a much more complete skeleton, was named and described by Stephen L. Brusatte and colleagues in 2009. Its relationships to other tyrannosaurid genera are unclear, with some evidence supporting a hypothesis that Alioramus is closely related to the contemporary species Tarbosaurus bataar. Alioramus were bipedal like all known theropods, and their sharp teeth indicate that they were carnivores. Known specimens were smaller than other tyrannosaurids like Tarbosaurus bataar and Tyrannosaurus rex, but their adult size is difficult to estimate since both species are known only from juvenile or sub-adult remains. The recent discovery of Qianzhousaurus indicates that it belongs to a distinct branch of tyrannosaur. The genus Alioramus is characterized by a row of five bony crests along the top of the snout, a greater number of teeth than any other genus of tyrannosaurid, and a lower skull than other tyrannosaurids. Alioramus remotus was estimated at 5 to 6 metres (16 to 20 ft) in length when originally described by Sergei Kurzanov in 1976. Kurzanov, however, did not correct for lengthening of the skull by deformation during fossilization, which may indicate a shorter overall body length for this individual. If this specimen is a juvenile, then adult Alioramus would have reached greater lengths, but no confirmed adult specimens are known. The skull of A. remotus was approximately 45 centimetres (18 in) long. In general, it is long and low, a shape typical of more basal tyrannosauroids and juveniles of larger tyrannosaurids. The premaxillary bones at the tip of the snout in Alioramus remotus have not been found, but are taller than wide in all tyrannosauroids for which they are known. The nasal bones are fused and ornamented with a row of five irregular bony crests that protrude upwards from the midline, where the nasal bones are sutured together. These crests all measure more than 1 centimetre (0.39 in) tall. At the back of the skull there is a protrusion, called the nuchal crest, arising from the fused parietal bones, a feature shared with all tyrannosaurids. In Alioramus, the nuchal crest is greatly thickened, similarly to Tarbosaurus and Tyrannosaurus. Like the rest of the skull, the lower jaw of Alioramus was long and slender, another possible juvenile characteristic. As in Tarbosaurus, a ridge on the outer surface of the angular bone of the lower jaw articulated with the rear of the dentary bone, locking the two bones together and removing much of the flexibility seen in other tyrannosaurids. Other tyrannosaurids had four premaxillary teeth, D-shaped in cross section, on each side. Including 16 or 17 in each maxilla, and 18 in each dentary, Alioramus had 76 or 78 teeth, more than any other tyrannosaurid. The braincase of A. altai was intermediate between the basal theropod and avialan conditions. The rest of the skeleton of Alioramus remotus is completely unknown except for three metatarsals (bones of the upper foot), but the discovery of A. altai, which is known from substantially more complete remains, has shed light on the anatomy of the genus. Paleontologists have long classified Alioramus within the superfamily Tyrannosauroidea, but because its remains were for many years poorly known, a more precise classification had remained elusive until the discovery of A. altai. A cladistic analysis published in 2003 found Alioramus could be further classified into the family Tyrannosauridae and the subfamily Tyrannosaurinae, alongside Tyrannosaurus, Tarbosaurus and Daspletosaurus. A 2004 study supported this result but suggested it was equally probable that Alioramus belonged outside the family Tyrannosauridae entirely, with its supposed juvenile characters actually reflecting a more basal position within Tyrannosauroidea. Another study omitted Alioramus altogether due to the only specimen's fragmentary nature. The discovery of A. altai in 2009 confirmed the placement of the genus within the Tyrannosaurinae. Tarbosaurus and Alioramus shared several skull features, including a locking mechanism in the lower jaw between the dentary and angular bones, and both lacked the prong of the nasal bones which connected to the lacrimal bones in all other tyrannosaurids except adult Daspletosaurus. The two genera may be closely related, representing an Asian branch of the Tyrannosauridae. Some specimens of Tarbosaurus have a row of bumps on the nasal bones like those of Alioramus, although much lower. The long and low shape of the only known Alioramus remotus skull indicated that it was immature when it died and might even have been a juvenile Tarbosaurus, which lived in the same time and place. The more prominent nasal crests and much higher tooth count of Alioramus, however, suggested it was a separate taxon, even if it is known only from juvenile remains, confirmed by the discovery of A. altai. Specimens identified as immature Tarbosaurus have the same tooth count as adults. The holotype (PIN 3141/1) of Alioramus is a partial skull associated with three metatarsals. A joint Soviet-Mongolian expedition to the Gobi Desert in the early 1970s found these remains at a locality known as Nogon-Tsav in the Mongolian province of Bayankhongor. Alioramus was named and described by Russian paleontologist Sergei Kurzanov in 1976. Its crests and low skull profile looked so different from other tyrannosaurids that Kurzanov believed his find was far removed from other members of the family. Accordingly, he gave it the generic name Alioramus, derived from the Latin alius ('other') and ramus ('branch'), and the specific name A. remotus, which means 'removed' in Latin. Alioramus is known from the holotypes of A. remotus and A. altai. The Beds of Nogon-Tsav are considered to be the same age as the Nemegt Formation. This geologic formation has never been dated radiometrically, but the fauna present in the fossil record indicate it was probably deposited during the Maastrichtian stage, at the end of the Late Cretaceous. The Maastrichtian stage in Mongolia, as preserved in the Nemegt Formation and at Nogon-Tsav, was characterized by a wetter and more humid climate compared with the semi-arid environment preserved in the earlier, underlying Barun Goyot and Djadochta Formations. Nemegt sediments preserve floodplains, large river channels and soil deposits, but caliche deposits indicate periodic droughts. This environment supported a more diverse and generally larger dinosaur fauna than in earlier times. Kurzanov reported that other theropods, including Tarbosaurus, ornithomimosaurs and therizinosaurs were discovered at the same locality, but these remains have never been reported in detail. If the Nogon Tsav fauna was similar to that of the Nemegt Formation, troodontid theropods, as well as pachycephalosaurs, ankylosaurids and hadrosaurs would also have been present. Titanosaurian sauropods were also potential prey for predators in the Nemegt. ( Read More ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9): ( Read More ) Размеры тела в сравнении с человеком:
Ископаемые останки и реплики (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, тираннозавриды, тираннозавроиды, целурозавры, ящеротазовые |
| Time | Event |
| 04:28 pm [industrialterro] [Link] |
Altirhinus Альтири́н (Altirhinus) — род птицетазовых динозавров из семейства игуанодонтид. Все окаменелости этого динозавра были найдены в 1981 году совместной Советско-Монгольской экспедицией в геологической формации Khukhtek, на территории Восточно-Гобийского аймака. Khukhtek датируется 125—100 миллионами лет, что относится к отложениям нижнего мела альбского—аптского ярусов. Известно несколько окаменелых скелетов. Типовым экземпляром является хорошо сохранившийся череп, а также некоторый посткраниальный материал, состоящий из частичных останков конечностей, плечевого и тазового поясов. Второй экземпляр включает частичный череп, связанный с рёбрами, фрагментарными позвонками, а также полными передними конечностями. Третий образец состоит из многих костей конечностей и 34 хвостовых позвонков оставшихся от меньшей особи. Невдалеке были обнаружены два скелета, вероятно, принадлежащим молодым особям этого вида. Первоначально останки альтирина были отнесены к виду Iguanodon orientalis, который был выделен в 1952 году. Однако, как показали дальнейшие исследования название Iguanodon orientalis было основано на чрезвычайно фрагментарных останках, которые вероятно на самом деле принадлежали Iguanodon bernissartensis. Останки, обнаруженные в Монголии, не соответствовали останкам игуанодона, поэтому в 1998 году они были выделены в отдельный род и вид. Altirhinus was herbivorous and bipedal when walking or running, but probably became quadrupedal when feeding from the ground. According to the original description, the entire body probably extended 26 feet (8 m) from snout to tail tip. In 2010 Gregory S. Paul estimated the length at 6.5 metres (21 ft), the weight at 1.1 tonnes. The skull alone is about 30 inches (760 mm) long, with a wide mouth and a distinctive tall arch on top of its snout, from which this dinosaur derives its name. All known specimens of Altirhinus were recovered in 1981 during collaborative expeditions organized by Soviet and Mongolian scientists, from the Khukhtek Formation in the Dornogovi Province of Mongolia. The Khukhtek was formed in the Aptian to Albian stages of the Early Cretaceous Period, which lasted from between 125 and 100 million years ago. Psittacosaurus and the primitive ankylosaurid Shamosaurus have also been found in these rocks. Several fossil specimens of different ages and sizes are known. The holotype, PIN 3386/8, is a skull, which is well preserved on the left side, as well as some postcranial material consisting of pieces of the hands, feet, shoulder and pelvic girdles. A more fragmentary skull was also recovered, associated with some ribs, fragmentary vertebrae, and a complete forelimb. A third specimen preserves many limb bones and a series of 34 tail vertebrae from a smaller individual. Two even smaller fragmentary skeletons, presumably of young individuals, were uncovered nearby. The remains of this animal were originally referred to the species Iguanodon orientalis, which was first described in 1952. However, I. orientalis has since been shown to be fragmentary, nondiagnostic, and virtually indistinguishable from the European I. bernissartensis (Norman, 1996). As no features of I. orientalis are shared exclusively with the 1981 specimens, which are clearly distinguishable from Iguanodon, a new name for those specimens was required. British paleontologist David B. Norman named them Altirhinus kurzanovi in 1998. The name was created from a Latin word, altus ("high") and a Greek word, ῥίς, rhis, genitive rhinos ("nose" or "snout"). There is one known species (A. kurzanovi), which honors Sergei Kurzanov, the influential Russian paleontologist who originally found the specimens in 1981. Altirhinus is definitely an advanced iguanodontian, just basal to the family Hadrosauridae, but there is little agreement on the arrangement of genera and species in this area of the ornithopod family tree. In the original description, it was included with Iguanodon and Ouranosaurus in a family Iguanodontidae (Horman, 1998). More recent analyses all find Altirhinus more derived than either of those two genera, but less than Protohadros, Probactrosaurus, and hadrosaurids (Head, 2001; Kobayashi & Azuma, 2003; Norman, 2004). The former two studies also place Eolambia between Altirhinus and hadrosaurids, while Norman's analysis finds that the two genera share a clade. Fukuisaurus is just basal to Altirhinus according to the only analysis in which the former has been included (Kobayashi & Azuma, 2003). Many aspects of Altirhinus' anatomy allow speculation on its behavior. As its forelimbs were roughly half the length of its hindlimbs, Altirhinus appears to have been primarily bipedal. However, its carpals (wrist bones) were thick and blocky, and the three middle fingers of its hand were wide, hyperextendable, and ended in hoof-like bones. This indicates that the forelimbs were also capable of supporting weight. Like many ornithopods, Altirhinus may have spent a significant amount of time in a quadrupedal position, perhaps while feeding. While the three middle digits of each forelimb (digits II, III, & IV) were very thick and probably weight bearing, the outside fingers (digits I & V) were modified in different ways. The first digit was a simple sharp spike, as seen in Iguanodon. Aside from defense, the thumb spike could possibly have also been used for breaking the shells of seeds or fruit. The fifth digit was somewhat opposable to the rest of the hand and may have been useful for grasping food. There is a large diastema, or gap, between the keratinous beak on the front of the mouth and the main chewing teeth in the side of the mouth, which would allow the two sections to work independently, so Altirhinus could crop with its beak while simultaneously chewing with its teeth. Many herbivorous mammals show a similar adaptation and can crop with their incisors without disturbing their chewing molars. Altirhinus was one of a number of advanced iguanodontians with snouts expanded outwards towards the end. This is quite possibly an example of convergent evolution with hadrosaurids, famous for their wide "duckbill" snouts. These adaptations are also paralleled in many living mammalian herbivores of different lineages. Modern cows, horses, and white rhinoceros all exhibit wide muzzles and all are grazing animals. Grazing most often occurs at ground level, and if the expanded muzzles of Altirhinus and other related species were an adaptation to grazing, this may also explain corresponding weight-bearing adaptations of the forelimbs in derived iguanodontians, in order to get the head closer to the ground. The characteristic arched snout of Altirhinus was formed primarily by the nasal bones, and a similar structure is seen on the snout of the Australian Muttaburrasaurus. Many different functions have been proposed for the nasal arch. It may have housed tissues to cool the blood, conserve water, or enhance the sense of smell. Alternatively, it may have facilitated communication through vocalization or visual display. As only two skulls have been located, it is entirely possible that the arched snout is only found in one gender, in which case it may have been used for sexual display, like in modern-day elephant seals. Гадрозавроиды (Hadrosauroidea) — клада или надсемейство птицетазовых динозавров, который включает в себя «утконосых» динозавров, или гадрозавридов, а также их близких родственников. Жили в меловом периоде на всех континентах планеты. Предполагается, что гадрозавроиды возникли в Азии и а потом широко расселились в другие части света. Самые ранние находки представителей данной группы относятся к аптскому веку раннего мелового периода, около 125 млн лет назад — это Probactrosaurus, живший на территории Китая, и Siamodon nimngami, найденный в Таиланде. Впервые название таксону дано Полом Серено в 1986 году в качестве подтаксона Iguanodontoidea. Ныне данная классификация не используется, и разные ученые предположительно относят гадрозавроидов либо к группе Hadrosauriformes (например Пол Серено в 1999 году и Годефруа (Godefroit) с коллегами в 2004 году), либо к группе Iguanodontia (например Ruiz-Omeñaca в 2006 году). Многие примитивные гадрозавроиды, такие как Ouranosaurus, традиционно включались в парафилетические группы игуанодонтидов. После кладистического анализа традиционных игуанодонтидов, данная группа в значительной степени распалась, и на данный момент включает в себя только игуанодона и его ближайших родственников. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, анкилополлексии, архозавроморфы, архозавры, гадрозавроиды, диапсиды, динозавроморфы, динозавры, игуанодонты, неорнитискии, орнитоподы, птицетазовые, цераподы |
















{kind=link}
_from_Mongolia.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
| Previous Day | 2015/04/24 [Archive] |
Next Day |