[Recent Entries][Archive][Friends][User Info]
Below are 7 entries, after skipping 40 most recent ones in the "Сообщество, посвящённое ра" journal:| August 6th, 2012 | |
|---|---|
| 12:11 pm [industrialterro] [Link] |
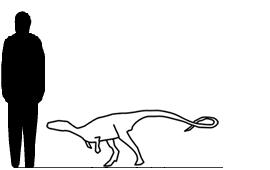
Chialingosaurus Chialingosaurus (meaning "Chialing Lizard") is a genus of stegosaur similar to Kentrosaurus from the Upper Shaximiao Formation, Late Jurassic beds in Sichuan Province in China. Its age makes it one of the oldest species of stegosaurs, living about 160 million years ago. Since it was an herbivore, scientists think that Chialingosaurus probably ate ferns and cycads, which were plentiful during the period when Chialingosaurus was alive. Its name is taken from the Chialing River in southern China. It may grow up to 4 metres (13 ft) long and weighing just 150 kg, much less than other later stegosaurs. Chialingosaurus was collected by the geologist Kuan in 1957, in Quxian County. The dinosaur was named by paleontologist C. C. Young two years later, although only very incomplete fossilized remains of Chialingosaurus have been found, and those fossil remains belong to a juvenile. In 1969, Rodney Steel suggested that Chialingosaurus might have actually been an early ancestor of other stegosaurs, but it is difficult to tell: the type specimen, Chialingosaurus kuani is known from only one partial skeleton, the original material having been supplemented in 1978 by Zhou of the Municipal Museum of Chongqing. Stegosauridae is a family of stegosauria, large thyreophorans. They lived longer than other Stegosaurs; while all Huayangosauridae and most of basal stegosaurs died out in Tithonian - Kimmeridgian, Stegosauridae survived till Middle Cretaceous. They are usually characterized by triangular plates on their back. These plates were not as hard as the plates of Huayangosauridae, so perhaps they were used only for display. However there are several exemptions: Dacentrurus, Lexovisaurus and Kentrosaurus have also spikes on their back. In the family tree of Thyreophora, Stegosauridae are right in the middle. Some paleontologists, propose a theory that they evolved from dinosaurs like Scelidosaurus. They state that in early stegosaurs, like Huayangosaurus, plates are relatively small, while in Stegosaurus, the most advanced member of the family, they are very large. Perhaps bony plates of stegosaurs evolved from scutes of Scelidosaurus or its relatives. Stegosaurids were usually large and powerful dinosaurs. Their front legs were shorter than their rear making them slow - moving dinosaurs. When Othniel Charles Marsh first found Stegosaurus, he portrayed him with very short front legs and neck. But new evidence says that they had relatively long these body parts. New discoveries suggest that in them was present sexual dimorphism. Stegosauria — группа птицетазовых динозавров входящая в кладу Thyreophora и живших с юрского по меловой периоды на всех континентах планеты, за исключением Антарктиды. Находки стегозавров Северной Америки ограничиваются единственной формацией Моррисона и представлены только 1 или 2 родами и несколькими видами. Все эти находки датируются кимериджем и титоном поздней юры. Разновидность даных динозавров в Англии, сравнительно более диверсифицированная, насчитывает около 6 родов, простирающихся от средней юры до раннего мела. К сожалению, имеющиеся окаменелости по большей части крайне разрознены и состоят из зубов, пластин и других костей, что делает их дальнейшую идентификацию довольно трудной и, в целом, сомнительной. Сюзанна Мейдмент и ее коллеги в своих работах 2008 года сделали общий пересмотр клады Stegosauria и признали только 2 валидных вида в Соединенном Королевстве: Dacentrurus armatus и Loricatosaurus priscus. Наиболее древние британские стегозавры относятся к батонскому веку (165-168 миллионов лет назад) средней юры и представлены несколькими изолированными костями, в том числе массивной правой бедренной кости молодой особи из формации Cornbrah в Оксфордшире (верхний батон), двумя неполными позвонками из формирования Sharp, Оксфордшир и двумя большими пластинами из формации Чиппинг Нортон в Глостершире. Все эти кости были отнесены к виду "Lexovisaurus" vetustus (= "Omosaurus" vetustus). Эти находки сделали семейство Stegosauridae старейшим в своей группе. Это значит, что, как минимум, один крупный вид стегозавров присутствовал в Европе в это время. Далее, из раннего келловейского века (161-165 миллионов лет назад) формации Оксфорд Клей был найден Lexovisaurus durobrivensis, известный по двум частичным скелетам. Мейдмент и др. в 2008 году определили его как недействительный вид, на том основании, что у него нет уникальной характеристики, присуствующей в голотипе образца. Вместо него они выделили новый род, Loricatosaurus, описаный по второму частичному скелету. Однако возможно эти два рода описывают одно и то же животное. Из формирования Coralline Oolite в Йоркшире, относящегося к среднему оксфордскому веку (156-161 миллионов лет назад), была выкопана плохо сохранившиеся бедренная кость молодой особи, которая была названа Omosaurus phillipsi (= Dacentrurus phillipsi). Нет никаких реальных признаков того, что это был стегозавр, и данное наименование считается nomen dubium (т.е. сомнительное название). В формировании Kimmeridge Clay в Уилтшире из нижнего кимериджского века (151-156 миллионов лет назад) был найден Dacentrurus armatus. Из Kimmeridge Clay также известно несколько костных шипов под названием "Omosaurus hastiger", которые могут принадлежать одному и тому же животному. Из нижнего мела в местности Wealden Beds, Сассекс валанжинского века (136-140 миллионов лет назад), найдена неполностью сохранившаяся челюсть Regnosaurus northamptoni, который также помечен как nomen dubium (Кумбс, 1971). Тем не менее, Барретт и Апчерч в 1995 году определили Regnosaurus как стегозавра и обнаружили, что этот динозавр принадлежал к старой линии стегозавров, которая включала также китайский род Huayangosaurus. Сомнительный вид Craterosaurus pottonensis, известный по одному неполному и плохо сохранившемуся спинному позвонку, был описан как стегозавр, но это, скорее всего, не так. Вид был найден в Бедфордшире, и вероятно, также относится к валанжинскому веку. Тиреофоры (Thyreophora — носители щита, от греч. θυρεος — большой продолговатый щит и греч. φορεω — нести) — один из двух подотрядов птицетазовых динозавров. Тиреофоры были травоядными динозаврами, которые жили с раннего юрского до конца мелового периода. Тиреофоры характеризуются наличием панцирных пластин, выстроенных продольными рядами вдоль тела. Тиреофоры включали в себя такие известные подотряды как анкилозавры и стегозавры, а также менее известные группы.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, птицетазовые, стегозавриды, стегозавры, тиреофоры |
| 11:42 am [industrialterro] [Link] |
Chaoyangsaurus Chaoyangsaurus ("Chaoyang lizard") was a marginocephalian dinosaur from the Late Jurassic of China (dated to between 150.8 and 145.5 million years ago). Chaoyangsaurus belonged to the Ceratopsia (Greek for "horned faces"), a group of primarily herbivorous dinosaurs with parrot-like beaks which thrived in North America and Asia during the Cretaceous Period (which ended roughly 65 million years ago). Chaoyangsaurus, like all ceratopsians, was primarily a herbivore. Chaoyangsaurus was found in the Chaoyang area of Liaoning Province in northeastern China. The specific name honours the Chinese paleontologist C. C. Young. Unlike many other dinosaurs, Chaoyangsaurus had been discussed in a number of sources before its official publication. As a result of this, several different spellings of its name have come and gone as nomina nuda ("naked names", names with no formal description behind them). The first name to see print was Chaoyoungosaurus, which appeared in the guidebook to a Japanese museum exhibit, and was the result of an incorrect transliteration. Zhao (1983) also used this spelling when he first discussed the species, so it is technically a nomen nudum. Two years later, Zhao again used this early spelling when he assigned a type specimen and species name, Chaoyoungosaurus liaosiensis (Zhao, 1985). According to Dong (1992), the name Chaoyoungosaurus had been officially described in a paper by Zhao and Cheng in 1983, but no cite for this supposed paper exists, and it is likely it was not properly published. Dong, in his 1992 paper on the subject, also emended the name to the "correct" spelling of Chaoyangosaurus (note the extra letter "o"). However, since this renaming was not accompanied by a formal description of the dinosaur, Chaoyangosaurus must also be considered a nomen nudum. It was not until 1999 that the dinosaur finally received an official name. Sereno (1999) used the name Chaoyangsaurus in an overview of ceratopsian taxonomy. Once again, that name was a nomen nudum. However, in December of that year, Cheng, Zhao, and Xu published an official description using the name Chaoyangsaurus youngi, and as the first name for this genus that is not a nomen nudum, it has official priority over all other spellings that have been used. Chaoyangsauridae is a family of dinosaurs. They were the first marginocephalian dinosaurs to appear on the planet, about 148 million years ago, in the Late Jurassic period. Members of this group had sharp beaks for snipping off leaves to eat, and a very small frill. There are disagreements that the chaoyangsaurids were part of the pachycephalosaurian group, ceratopsian group, or a different ancestral group. Two dinosaur genera, Chaoyangsaurus and Xuanhuaceratops, are included in Chaoyangsauridae. Both animals are more primitive (or basal) than Psittacosaurus, an early ceratopsian. Цера́топсы, рогатые динозавры (Ceratopsia, от греч. κέρας — «рог», ωψ — «лицо») — группа (инфраотряд) травоядных птицетазовых динозавров позднего юрского и мелового периодов, обладавших характерным крючковатым клювом. Окаменелые останки обнаружены в Северной Америке, Азии и Европе. Из наиболее известных представителей — цератопс и трицератопс. Происхождение и эволюция цератопсов окончательно не ясны. Самым ранним цератопсом считается открытый в Китае в 2005 году динозавр Иньлун (Yinlong), чьи останки датируются поздним юрским периодом (161—156 млн лет назад), в результате чего предполагается их азиатское происхождение в юрском периоде. Также остается открытым вопрос и о появлении крупных рогатых динозавров в позднем меле на территории Северной Америки. Находки палеонтологов в 2010 году на территории Венгрии дали доказательство тому, что в среднем меловом периоде цератопсы (Ajkaceratops) обитали на территории современной Европы, которая в те времена представляла собой разрозненные острова. Учёные предполагают, что данная разновидность мигрировала из Азии, двигаясь от острова к острову. Marginocephalia ("fringed heads") is a clade of ornithischian dinosaurs that includes the thick-skulled pachycephalosaurids, and horned ceratopsians. They were all herbivores, walking on two or four legs, and are characterized by a bony ridge or frill at the back of the skull. The clade evolved in the Jurassic period, and became common in the Upper Cretaceous.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, птицетазовые, цераподы, цератопсы, чаоянгзавриды |
| August 3rd, 2012 | |
| 02:30 pm [industrialterro] [Link] |
Camptosaurus Camptosaurus ( On September 4, 1879 William Harlow Reed in Albany County, Wyoming found the remains of a small euornithopod. That same year Professor Othniel Charles Marsh described and named the find as Camptonotus, or "flexible back", from Greek κάμπτω, "to bend" and νῶτον, "back", in reference to the presumed flexibility of the sacral vertebrae. The holotype was YPM 1877, a partial skeleton. The genus was renamed Camptosaurus by him in 1885 because the original name was already in use for a cricket. In 1879, Marsh named C. dispar (type species of the genus) for material he received from his collectors at Quarry 13 near Como Bluff, Wyoming in the Morrison Formation and C. amplus based on the holotype YPM 1879, a foot found by Arthur Lakes at Quarry 1A. The foot was later shown to have belonged to Allosaurus. Throughout the 1880 and 1890s, he continued to receive specimens from Quarry 13 and in 1894 named two additional species: C. medius and C. nanus, based in part on size. Charles W. Gilmore named two additional species, C. browni and C. depressus in his 1909 redescription of the Marsh specimens. In the Morrison Formation, Camptosaurus fossils are present in stratigraphic zones 2-6. Then in 1980, Peter Galton and H.P. Powell in their redescription of C. prestwichi (see following), considered C. nanus, C. medius and C. browni to be different growth stages or different gender of the larger C. dispar, and therefore only C. dispar was a valid species. They also considered a skull, YPM 1887, in 1886 referred to C. amplus by Marsh, later confirmed by Gilmore, to belong to C. dispar as well. Gilmore had used this skull to describe the skull of Camptosaurus, but the specimen was recently shown by Brill and Carpenter not to belong to Camptosaurus. In 2007, they put it into its own genus and species, Theiophytalia kerri. Camptosaurus depressus was recovered from the Lakota Formation near the town of Hot Springs, South Dakota. It was described by Charles Gilmore in 1909 based on the holotype and only known specimen USNM 4753, a fragmentary postcranium, by the "narrowness or depressed nature of the ilia". Carpenter and Wilson (2008) referred this species to Planicoxa, as P. depressa, on the basis of similarities between its ilium and the holotype ilium of Planicoxa venenica. However, McDonald and colleagues (2010), and McDonald (2011) found that the horizontal postacetabular process of C. depressus is more likely a product of distortion. Therefore, McDonald put it into its own genus, Osmakasaurus. An additional species, Camptosaurus aphanoecetes, was named by Carpenter and Wilson in 2008 for specimens from Dinosaur National Monument. It differs from C. dispar in the lower jaw, shorter neck vertebrae, and straighter ischium ending in a small "foot" among other features. An analysis by Andrew McDonald and colleagues in 2010 suggested that like Cumnoria, C. aphanoecetes is actually more closely related to more advanced iguanodonts (Styracosterna). It has been moved to the new genus Uteodon. Due to the separate status of Uteodon it has become problematic which material from the Morrison belongs to Camptosaurus. The specimens with certainty belonging to Camptosaurus dispar, from Quarry 13, have been recovered from very deep layers, probably dating to the Callovian-Oxfordian. The largest fragments from later strata indicate adult individuals more than 7.9 metres (26 ft) long, and 2 metres (6.6 ft) at the hips. Based on studies of other iguanodonts, scientists believe they may have been able to achieve running speeds of 25 km per hour (15 mph). The Quarry 13 individuals are smaller though. They have been described as reaching 6 metres (19.7 feet) in length and 785 – 874 kg in weight. In 2010 Gregory S. Paul gave an even lower estimate: a length of five metres and a weight of half a tonne. Camptosaurus is a relatively heavily built form, with robust hindlimbs and broad feet, still having four toes. The wrists are strongly ossified allowing the animal to rest on its arms while feeding from low vegetation. The fingers can be spread, not being bound together in a pad, and the thumb carries a small spike. Because Marsh and Gilmore based themselves on the skull of Theiophytalia, many earlier reconstructions of the head of Camptosaurus have been incorrect, showing a much too rectangular profile. The skull was in fact triangular with a pointed snout, equipped with a beak. Its teeth were more tightly packed in the jaw compared to other Morrison euornithopods. Museum curator John Foster describes them as having "thick median ridges on their lateral sides and denticles along their edges," these features were similar to, but "more fully developed" than those in Dryosaurus. Camptosaurus teeth frequently exhibit extensive wear, which indicates that individuals in the genus had a diet of relatively tough vegetation. A tiny 9 inch fossilized embryo, referred to Camptosaurus, was retrieved from Morrison Formation strata at Dinosaur National Monument in Utah. Marsh in 1885 assigned Camptosaurus to a family of its own: the Camptosauridae. Alternatively some authors considered it an early member of the Iguanodontidae. Modern phylogenetics has made Camptosaurus by definition part of the clade Ankylopollexia, of which group it would then be a basal member. This would mean that the genus is closely related to the ancestor of later iguanodontid and hadrosaurid dinosaurs and was more derived than contemporaries such as Dryosaurus, Drinker, and Othnielosaurus. Нижняя челюсть камптозавра содержала несколько рядов зубов с неровными краями, способных без труда срезать и измельчить растительный материал. Края зубов были покрыты множеством мелких острых выступов, легко срезавших листья и ветки. Поскольку растения были очень жесткими, режущие кромки зубов стирались практически полностью. Длинное рыло камптозавра заканчивалось роговым клювом, которым он крепко захватывал и удерживал растения. Вероятнее всего камптозавр имел бочкообразное тело, поскольку ему требовался большой желудок, чтобы переваривать огромный обьем листьев, побегов и веток. У камптозавра были длинные, сильные задние лапы и укороченные передние, поэтому он, скорее всего, передвигался на двух конечностях. Хотя передние лапы камптозавра были слабыми, для того чтобы выдержать значительный вес его теда, кости стопы и кисти были устроены так, что при необходимости камптозавр мог опираться и на все четыре конечности. Это позволяло срывать растения растущие близко к земле. Таким образом динозавр мог легко наклоняться вниз, используя на короткое время передние лапы как подпорки. Камптозавр иногда поднимался на задних лапах, чтобы сорвать сочные молодые побеги. Длинный и тяжелый хвост камптозавра служил в таком случае противовесом. Связки представляют собой прочные, похожие на струну, ткани, соединяющие мышцы с костями. Они имеются в любой части тела, в том числе и на спине, усиливая позвоночник. У большинства животных связки состоят из мягкого, но прочного материала, называемого коллагеном. Связки молодого камптозавра сначала состояли из коллагена, но по мере роста животного коллаген заменялся костным материалом. У взрослого камптозавра связки выглядили как сеть очень длинных тонких костей. Эти кости не только укрепляли позвоночник, но и помогали камптозавру удерживать равновесие, придавая прямую и ровную осанку. Имея длинные задние и короткие передние конечности, капмтозавр ходил только на задних лапах. Он был достаточно тяжелым и вероятно, не мог быстро бегать. В отличие от игуанодона, у камптозавра шип на большом пальце кисти отсутствовал. Пятипалые кисти и четырехпалые ступни заканчивались не когтями, а скорее копытцами. Камптозавры жили стадами и, возможно, уходили на новые места, когда истощались запасы корма. Палеонтологи считают, что камптозавры обладали своеобразными защечными мешками, в которые они набивали большие порции листьев и веточек. Там этот комок пережевывался короткими острыми зубами, расположенными в несколько рядов. Мощные четырехпалые задние ноги помогали уйти от преследователя. Опираясь на них, камптозавр длинным языком собирал листья с деревьев и саговников, объедал низкорослые растения. Хотя в передней части клювовидной пасти камптозавра не было зубов, он мог пережёвывать растения многочисленными зубами, которые росли в глубине рта. Останки этого динозавра обнаружены на территории Европы и Северной Америки. Ankylopollexia is a clade of herbivorous iguanodontian dinosaurs that lived from the Late Jurassic to Late Cretaceous in all continents. Some members include Camptosaurus, Iguanodon, Ouranosaurus, and the hadrosaurids or "duck-billed dinosaurs". The Ankylopollexia was named by Paul Sereno in 1986 to denote the group within the Iguanodontia that includes the Camptosauridae and the Styracosterna. The name means "stiff thumbs", referring to a possible synapomorphy, possessing a conical thumb spine. Ankylopollexians were generally large animals, and some (such as Shantungosaurus, which measured up to 50 ft (15 m) in length and weighed up to 8 tons) equaled the largest carnivorous dinosaurs in size. The first definition of Ankylopollexia was given by Sereno in 1998. It was defined as the node-based taxon consisting of the last common ancestor of Camptosaurus, Parasaurolophus and all his descendants. Sereno (2005) improved this definition, using the type species, Camptosaurus dispar (Marsh 1879) and Parasaurolophus walkeri (Parks 1922), instead of the genera. Many ankylopollexians have not yet been included in a large phylogenetic analyses, or are too fragmentary to place confidently. These include Bihariosaurus, Delapparentia, Dollodon, Draconyx, Hypselospinus, Lurdusaurus, Osmakasaurus, Owenodon, Planicoxa, Proplanicoxa and Sellacoxa. The simplified cladogram below follows an analysis by Andrew McDonald and colleagues, published in November 2010 with information from McDonald, 2011. Орнитоподы (Ornithopoda, от др.-греч. ὄρνις, род. п. ὄρνιθος — птица и др.-греч. πούς — «нога») — инфраотряд птицетазовых травоядных динозавров. Жили от позднего юрского периода до конца мелового периода. Многие из них имели крупные размеры, игуанодон, встав на дыбы, достигал 10 (в некоторых случаях даже 13) метров. Передние лапы у них обычно были значительно более короткими (в 1,5—2 раза) и не столь сильными, как задние. Поэтому ранее считалось, что они передвигались исключительно на задних конечностях; согласно последним исследованиям, использовали при ходьбе также и передние лапы. Зубы срастались по всей длине челюсти, а про вертикали состояли из нескольких коронок, заменявшихся новыми по мере стачивания. Игуанодоны — наиболее известные и ранее всего открытые динозавры этой группы. Гадрозавры и родственные им виды за своеобразную форму морды, напоминающую клюв, получили прозвание «утконосых». Они были меньше игуанодонов (3—4 м). Цераподы (Cerapoda) — один из двух подотрядов или групп птицетазовых динозавров. Цераподы были травоядными динозаврами, которые жили на всех континентах. Название таксону было дано Полом Серено в 1986 году (таксон был указан как "Nanorder"). Ныне первоначальная классификация была пересмотрена и не сохранилась. Было высказано мнение о монофилетичности группы Батлером и его коллегами в 2008 году, а также Салливаном в 2009 году. Некоторые ученые относят Micropachycephalosaurus hongtuyanensis в семейство пахицефалозавров. Батлер и Ци (2009) считают его членом клады Ceropoda, потому что голотипу не хватает "однозначной синапоморфии пахицефалозавра", в то время как Салливан (2006) считает, что данный таксон является вовсе nomen dubium. Голотип Micropachycephalosaurus hongtuyanensis, в котором отсутствуют череп материала, считался взрослой особью Перл и др.. (1982), но недавно это было пересмотрено, и теперь он определяется как несовершеннолетняя особь (Батлер и Ци, 2010).
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие рептилии, Юра, авеметатарзалии, анкилополлексии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, игуанодонты, неорнитискии, орнитоподы, птицетазовые, цераподы |
| May 29th, 2012 | |
| 07:03 pm [industrialterro] [Link] |
Agilisaurus Агилизавр (Agilisaurus) — род вымерших птицетазовых динозавров из группы Neornithischia, живших в среднем юрском периоде (около 171,6—167,7 миллионов лет назад), на территории нынешней Азии. Окаменелости были найдены в провинции Сычуань в Китае. Впервые описан палеонтологом Пенгом в 1990 году. Представлен одним видом — Agilisaurus louderbacki. Хотя агилизавр известен по почти полному скелету, его фактическое положение в эволюционной иерархии динозавров до сих пор остается неясным. Пол Барретт из Лондонского музея естественной истории, считает, что он слишком примитивен, чтобы отнести его к традиционным орнитоподам. Возможно, это фаброзаурид, один из очень примитивных орнитопод, еще не имевших защечных пазух, но не исключено, что он является представителем совершенно нового семейства. Череп агилизавра маленький, с большими глазами, его зубы имеют форму листьев. Зубы крупные, в передней части они заострены. Задние конечности гораздо длиннее передних. Бедренная кость кажется особенно короткой по сравнению с остальными костями ног: признак быстро бегающего животного с легкой стопой и мускулами, сосредоточенными у бедра. Длинный хвост использовался для баланса при беге. Agilisaurus ('agile lizard') is a genus of ornithischian dinosaur from the Middle Jurassic Period of what is now eastern Asia. The name is derived from the Latin agilis meaning 'agile' and the Greek sauros meaning 'lizard', and refers to the agility suggested by its lightweight skeleton and long legs. Its tibia (lower leg bone) was longer than its femur (upper leg bone), which indicates that it was an extremely fast bipedal runner, using its long tail for balance, although it may have walked on all fours when browsing for food. It was a small herbivore, about 1.2 meters (4 feet) in length, and like all ornithischians, it had a beak-like structure on the ends of both upper and lower jaws to help it crop plant material. Despite its completeness, Agilisaurus has been placed in many different positions in the ornithischian family tree. It was originally placed in the family Fabrosauridae, which is no longer considered valid by most paleontologists (Peng 1990). Several recent studies, including cladistic analyses, find Agilisaurus to be the most basal member of the group Euornithopoda, which includes all ornithopods more derived than the family Heterodontosauridae (Weishampel et al. 2003; Norman et al. 2004). However, heterodontosaurs are not universally considered to be ornithopods and have been considered more closely related to the suborder Marginocephalia, which includes ceratopsians and pachycephalosaurs. In one recent cladistic analysis, Agilisaurus was found in a position basal to heterodontosaurs in the branch leading to Marginocephalia (Xu et al. 2006). Agilisaurus has been recovered in other positions as well, including as an ornithischian basal to both ornithopods and marginocephalians (Barrett et al. 2005; Butler 2005; Butler et al. 2008). Comparisons between the scleral rings of Agilisaurus and modern birds and reptiles suggest that it may have been diurnal, unlike larger herbivorous dinosaurs that were inferred to be cathemeral, active throughout the day at short intervals. Neornithischia ("new ornithischians") is a clade of the dinosaur order Ornithischia. They are the sister group of the Thyreophora within the clade Genasauria. Neornithischians are united by having a thicker layer of asymmetrical enamel on the inside of their lower teeth. The teeth wore unevenly with chewing and developed sharp ridges that allowed neornithischians to break down tougher plant food than other dinosaurs.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, неорнитискии, птицетазовые |
| May 28th, 2012 | |
| 08:02 pm [industrialterro] [Link] |
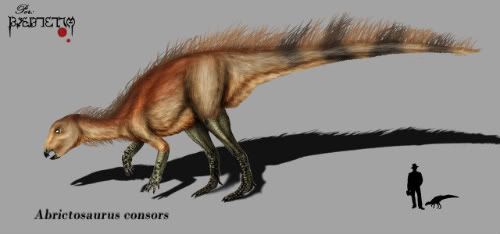
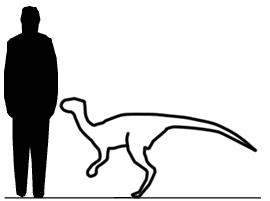
Abrictosaurus Абриктозавр (Abrictosaurus; букв. «бодрствующий ящер») — род динозавров из группы гетеродонтозавридов, живших в раннем юрском периоде на территории нынешней Южной Африки. Это был мелкий двуногий травоядный динозавр, около 1 метра в длину, и весом менее 45 килограмм. Этот динозавр известен только по двум скелетам, найденным в районе Цгачас-Нек в Лесото и в Капской провинции Южной Африки. Свита, в которой были обнаружены скелеты, которые относится к геттангскому и синемюрскому ярусам раннего юрского периода, около 200—190 миллионов лет назад. Имя «широко зевающий ящер» возникло в результате спора между палеонтологами. Тони Талборн предположил, что гетеродонтозавриды проводили значительную часть засушливого периода в спячке, как это делают некоторые современные животные. Дж. А. Хопсон выдвинул свою гипотезу, основанную на изучении расположения зубов, согласно которой животное было активно круглый год, и дал новому динозавру подходящее имя. Наши представления об абриктозавре основаны на двух черепах и нескольких фрагментах скелета. Черепа почти идентичны черепам гетеродонтозавра с таким же расположением зубов, за исключением отсутствующих впереди бивней. Возможно, найденные черепа принадлежали самкам гетеродонтозавра, а самцы имели бивни и использовали их для демонстрации силы, как это делают многие современные животные. С другой стороны, многие современные группы животных, такие как свиньи, имеют выступающие бивни у некоторых родов, в то время как у других родов, относящихся к этой группе, они отсутствуют. Heterodontosaurids like Abrictosaurus were small, early ornithischians, named for their markedly heterodont dentition. They are best known for the large, canine-like tusks (often called caniniforms) in both upper and lower jaws. There were no teeth in the front of the jaws, where a hard beak was used to crop vegetation. There were three premaxillary teeth, with the first two small and conical and the third enlarged to form the upper caniniform, counterpart to the even larger lower caniniform, which was the first dentary tooth. In the upper jaw, a large gap (or diastema) accommodated the lower caniniform tooth and separated the premaxillary teeth from the wider chewing teeth of the maxilla. Similar teeth lined the remainder of the lower jaw. Abrictosaurus is usually considered the most basal member of the family Heterodontosauridae. Lycorhinus and Heterodontosaurus both had high-crowned cheek teeth, which overlapped each other in the jaw, forming a continuous chewing surface analogous to those of Cretaceous hadrosaurids. Abrictosaurus had more widely-separated cheek teeth, with lower crowns, more similar to other early ornithischians. It has been suggested that Abrictosaurus lacked tusks and that this is another primitive feature. However, caniniforms were clearly present on one of the two specimens of Abrictosaurus. The upper caniniform measured 10.5 millimeters (0.4 inches) high, while the lower reached 17 mm (0.67 in). These caniniforms were serrated only on the anterior surface, unlike those of Lycorhinus and Heterodontosaurus, which were serrated on both anterior and posterior edges. Abrictosaurus also had smaller, less powerful forelimbs than Heterodontosaurus and one fewer phalanx bone in both the fourth and fifth digits of the forelimb. Гетеродонтозавриды (Heterodontosauridae), так же Lycorhinidae — семейство ранних птицетазовых динозавров, которые, вероятно являются одним из самых базальных (примитивных) членов группы. Хотя их окаменелости встречаются довольно редко, жили они почти по всему миру. Останки их обнаружены в Африке, Азии, Европе, Северной и Южной Америке. Гетеродонтозавры появляются в палеонтологической летописи в начале юрского периода (около 199 млн лет назад). Известно также несколько поздних видов из раннего мела (около 112 млн лет назад). Наиболее продвинутые члены семейства известны по находкам из Южной Африки и датированы раннеюрским периодом, в то время как наиболее примитивные и наименее специализированные члены группы, например Tianyulong, относятся к раннему меловому периоду. Среди ученых нет единого мнения, с чем это связано. Гетеродонтозавры являются редкой в отношении ископаемых группой, и мало что было известно об них до 2000 года. Это связано не только с дефицитом их останков, но и с большим пробелом в летописи окаменелостей ранних птицетазовых (не было достаточного количества находок других представителей отряда для сравнения). Перед 2000 годом достоверно было известно о существовании только трёх видов - Heterodontosaurus tucki, Abrictosaurus consors и Lycorhinus angustidens, все из ранней юры Южной Африки, с добавлением некоторых сомнительных таксонов, Lanasaurus, Geranosaurus (оба из Южной Африки) и загадочного Echinodon becklesii из раннего мела Англии, который известнен по фрагменту челюсти с зубами, которые могут принадлежать или не принадлежать гетеродонтузавру (Norman & Barrett, 2002). Недавние исследования показали, что Echinodon находится вне клады Heterodontosauridae. Однако все изменилось довольно быстро в течение последующих несколько лет. Все предыдущие неоспариваемые гетеродонтозавриды были из ранней юры Южной Африки. Открытия в новом тысячелетии значительно расширили временной и географический диапазон для группы, охватывая поздний триас Южной Америки (Баэс и Марсикано, 2001), ранний мел Китая (Чжэн в соавт., 2009) и верхнюю юру из Северной Америки (Батлер в соавт., 2010). Крупнейший представитель данного семейства Lycorhinus angustidens из ранней юры Южной Африки в основном известный по ископаемым останкам зубов, по оценкам достигал в длину всего 2 метра, в то время как Fruitadens haagarorum из верхней юры Колорадо, был около 65-75 см и являлся самым маленьким среди изученых птицетазовых динозавров на момент его открытия. На основе гетеродонтозавра Tianyulong из Китая, можно предположить, что эти динозавры были покрыты примитивными перьями, что указывает на раннее появление теплоизолирующих покровов у различных, эволюционно не связанных друг с другом, групп динозавров.
Tags: Вымершие рептилии, Юра, авеметатарзалии, архозавроморфы, архозавры, гетеродонтозавры, диапсиды, динозавроморфы, динозавры, птицетазовые |
| March 24th, 2012 | |
| 01:19 pm [industrialterro] [Link] |
Pisanosaurus Pisanosaurus (meaning "Pisano lizard") is a genus of primitive ornithischian dinosaur from the Late Triassic of what is now South America. It was a bipedal herbivore described by Argentine paleontologist Rodolfo Casamiquela in 1967. Only one species, the type, Pisanosaurus mertii, is known, based on a single partial skeleton. The fossils were discovered in Argentina's Late Triassic Ischigualasto Formation, which dates to about 228 to 216.5 million years ago. The exact classification of Pisanosaurus has been the topic of debate by scientists for over 40 years; the current consensus is that Pisanosaurus is the oldest known ornithischian, part of a diverse group of dinosaurs which lived during nearly the entire span of the Mesozoic Era. Based on the known fossil elements, Pisanosaurus was a small, lightly built dinosaur approximately 1 m (3 ft 3 in) in length. Its weight was between 2,27–9,1 kg (5–20 lb). These estimates vary due to the incompleteness of the fossil. The tail of Pisanosaurus has been reconstructed as being as long as the rest of the body, based on other early ornithischians, but as a tail has not been recovered, this is speculative. It was bipedal and, like all other known ornithischians, was probably herbivorous. Pisanosaurus mertii was described by Argentine paleontologist Rodolfo Casamiquela in 1967. The name Pisanosaurus honors Juan A. Pisano, an Argentine paleontologist, while saurus is derived from the Greek σαυρος, meaning "lizard". Pisanosaurus is known from a single fragmented skeleton found in Argentina. It is based on a specimen given the designation PVL 2577, which was discovered in the Ischigualasto Formation. Pisanosaurus is very basal within Ornithischia; the postcrania seem to lack any good ornithischian synapomorphy; it was even suggested by Paul Sereno in 1991 that the fossil is a chimera. However, recent studies suggest that the fossils belong to a single specimen. Pisanosaurus has been classified either as a heterodontosaurid or as the earliest known ornithischian. A 2008 study placed Pisanosaurus outside of (and more basal than) Heterodontosauridae. In this study, Pisanosaurus is the earliest and most primitive ornithischian. Pisanosaurus is the type genus of the Pisanosauridae, a family erected by Casamiquela in the same paper which named Pisanosaurus. The Pisanosauridae family has fallen into disuse, as a 1976 study considered the group synonymous with the already named Heterodontosauridae.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, птицетазовые |
| March 17th, 2012 | |
| 06:09 pm [industrialterro] [Link] |
Eocursor Эокурсор (Eocursor; букв. «ранний бегун») — род вымерших птицетазовых динозавров, живших в позднем триассовом периоде (около 216,5 - 203,6 миллионов лет назад), на территории нынешней Южной Африки. Впервые описан палеонтологом Ричардом Батлером (Richard Butler) и его коллегами в 2007 году. Представлен одним видом - Eocursor parvus. Название рода Eocursor образовано от греческого слова «eos» – «рассвет» или «начало» и латинского «cursor» – «бегун». Видовое название образовано от латинкого «parvus» – «маленький». Окамнелые останки Eocursor были найдены в 1993 году на территории фермы Damplaats (55), близ города Ladybrand, ЮАР, в геологической формации Lower Elliot относящейся к норианскому ярусу. Голотип SAM-PK-K8025 состоит из частичного скелета. Реконструированый внешний вид, а также описание динозавра было представлено только в 2007 году палеонтологами R. Butler, R. Smith и D. Norman. Принадлежит к плохо изученным на сегодняшний день ранним птицетазовым динозаврам (Ornithischia). По внешнему виду сильно отличается от общепринятого облика рептилий данного отряда. Искомпаемые останки включают в себя кости черепа, конечностей, таза и позвоночника, и являются наиболее полными известными останками скелета птицетазового динозавра триасового периода на территории Южной Африки. В длину Eocursor parvus достигал 1 метра, а высота не превышала 30 см. Листообразные зубы ящера предназначались для пережевывания исключительно растительной пищи. Верхние конечности, обладавшие большими, несоизмеримыми с телом, размерами, были очень удобными для хватания. А строение костей ящера и их небольшой вес свидетельствует о том, что животное могло быстро и проворно перемещаться. Птицетазовые (Ornithischia) — один из двух отрядов динозавров. Несмотря на своё название, не они, а ящеротазовые динозавры по одной из версий были предками птиц. Были травоядными (за исключением ранних всеядных видов), меньших размеров, чем ящеротазовые зауроподы. Кроме самых примитивных, имеют приспособления к питанию растительной пищей: сложный кишечник и щёки. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Таз имел четырёхлучевое строение. Лобковая кость имела вырост, направленный назад вдоль седалищной кости (у ящеротазовых кости были практически перпендикулярны). Лобковая кость часто образует новый вырост, направленный вперёд параллельно позвоночнику. Подобное строение таза развивалась у динозавров независимо трижды: у птицетазовых, у надсемейства Therizinosauroidea и семейства Dromaeosauridae. Последнее и является наиболее вероятным предком птиц. У некоторых видов (Hypsilophodon, Talenkauen и Thescelosaurus) имеются межреберные костные пластины в передней части ребер. Некоторые исследователи предполагают, что эти пластины выполняли биомеханические функции, помогали в дыхании либо были предназначены для обеспечения повышенной жесткости передней части грудной клетки, в то время как другие полагают, что они служили для оборонительных функций. Анализ имеющихся у ученых останков птицетазовых динозавров, позволил им определить, что ящеры данного отряда впервые появились в позднем триасе, примерно 220 млн лет назад и вначале были крайне малочисленны. Этот отряд динозавров начал бурно эволюционировать только в последующий юрский период (208–144 млн лет назад), все больше распространяясь по планете и занимая экологические ниши после исчезновения иных травоядных существ. Ранее птицетазовые делились на 4 или 6 подотрядов, включая тиреофоров (иначе (стегозавров), рогатых динозавров (цератопсов), анкилозавров (панцирных динозавров), цераподов (известнейшие представителей которых — игуанодоны) и панцироголовых пахицелозавров. В настоящее время они подразделяются на два подотряда-клады: тиреофоров и цераподов. Первые включают инфраотряды стегозавров и анкилозавров, вторые — рогатых динозавров и пахицефалозавров (со своей стороны нередко объединяемых в кладу или подотряд маргиноцефалов), и орнитоподов (включая игуанодонов, утконосых гадрозавров и др.)
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, птицетазовые |



































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}