[Recent Entries][Archive][Friends][User Info]
Below are the 20 most recent journal entries recorded in the "Сообщество, посвящённое ра" journal:| October 31st, 2011 | |
|---|---|
| 08:24 pm [industrialterro] [Link] |
Tetraceratops Тетрацератопс (Tetraceratops insignis) — синапсид неясного систематического положения. Вероятнее всего, относится к пеликозаврам (может быть родственником примитивных сфенакодонтов-хаптодусов). С 1990 года считался самым примитивным представителем терапсид, сейчас это вызывает сомнения. Единственный череп обнаружен Чарльзом Штернбергом в 1897 году в раннепермских слоях Байлора, в Техасе (формация Арройо, около 280 млн. л.н.). Описан У.Д. Мэтью в 1908 г. Первоначально считался пеликозавром из группы эотирид. Череп длиной около 10 см, очень короткий и высокий. Мощные передние «резцы», затем идет диастема, присутствует пара крупных верхних «клыков». На нижней челюсти клыков нет (есть увеличенные передние зубы, остальные зубы примерно одинакового размера). Самое интересное — наличие «рогов». Это невысокие выросты черепа с отпечатками сосудов, напоминающие «рога» терапсид — бурнетиид. Две пары «рогов» находятся на верхней поверхности черепа (пара — на предчелюстных костях, пара — на предлобных). Еще пара «рогов» обнаружена не так давно при дополнительном препарировании черепа — они растут из угловых отростков нижней челюсти. Таким образом, всего «рогов» было шесть. Скелет неизвестен, но «парус» имелся вряд ли. Был мелким хищником, вероятно, обитавшим на равнинах вдали от водоемов. Не исключено, что описанный Э. Олсоном в 1962 году из более поздних пермских отложений (Сан-Анжело, около 275 - 277 млн .л.н.) «фтинозухид» горгодон (Gorgodon minutus) — тоже тетрацератопс.
Tags: Вымершие синапсиды, Пермь, пеликозавры, терапсиды |
| 08:09 pm [industrialterro] [Link] |
Steppesaurus Степпезавр (Steppesaurus gurleyi) – пеликозавр неясного систематического положения, вероятнее всего, сфенакодонт. Происходит из «среднепермских» отложений (свита Флоуэр-пот, кунгурская эпоха) Техаса. Описан Э. Олсоном и Дж. Бирбауэром в 1953 году по фрагментарным остаткам. Согласно описанию Олсона: «Максиллярные зубы длинные, очень сильно сжаты с боков, с пильчатым передним и задним краями. Лингвальные поверхности коронок слабо вогнутые. Впереди первого зуба челюстной кости имеется хорошо развитый уступ челюстного края. Зубы постепенно уменьшаются спереди назад. Нижняя челюсть слабая, с рядом сильно уплощенных латерально зубов. Замещение щечных зубов одновременное; замещающие зубы чередуются с альвеолами зубов предшествующей генерации». Размеры крупные – длина челюсти до 59 см, морда длинная, чем напоминает эотитанозухий. Олсон первоначально указывал на наличие у этого рода удлиненных остистых отростков, как у диметродона, хотя в голотип входят только максилла, часть нижней челюсти и разрозненные зубы. Позвонки, как позже выяснилось, принадлежали Dimetrodon angelensis. В 1962 году Э.Олсон предположил, что степпезавр принадлежит к примитивным терапсидам. Переизучение материалов другими исследователями в 1986 и 1995 годах показало, что степпезавр и прочие «эотериодонты» Олсона – в действительности пеликозавры. Обычно пишут, что все описанные Олсоном терапсиды в реальности основаны на фрагментарных остатках казеид. В частности, считавшийся близкородственным степпезавру ноксозавр (Knoxosaurus niteckii) в настоящее время фигурирует в списках казеид. Тем не менее, степпезавр остается в списках сфенакодонтов. Не вполне понятно, существовал ли степпезавр как отдельный род или это просто неверно описанные остатки крупного диметродона – например, Dimetrodon angelensis, жившего примерно в ту же эпоху в тех же местах. Отличие степпезавра от других сфенакодонтов, указанное Олсоном – необычный характер смены зубов (Олсон писал, что степпезавр "не имеет никакого сходства с диметродоном"). Не ясно, может ли такой характер смены зубов соответствовать, например, казеидам, но по размерам челюстей степпезавр превосходит любых, даже самых крупных казеид. Судя по фотографии из работы Э. Олсона, максилла степпезавра собрана из осколков и включает большое количество гипса. С другой стороны, присутствие в этих же отложениях крупного вида диметродона исключает возможность наличия второго сфенакодонта таких же размеров, хотя степпезавр, по-видимому, характерен для более поздних горизонтов свиты Флоуэр-пот. Степпезавр остается загадкой палеонтологии до появления новых находок.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 07:51 pm [industrialterro] [Link] |
Varanops Varanops is an extinct genus of Early Permian varanopid synapsid known from Texas and Oklahoma of the United States. It was first named by Samuel Wendell Williston in 1911 as a second species of Varanosaurus, Varanosaurus brevirostris. In 1914, Samuel W. Williston reassigned it to its own genus and the type species is Varanops brevirostris. V. brevirostris is known from the holotype FMNH UC 644, a three-dimensionally preserved nearly complete and articulated skeleton including an nearly complete skull and mandibles. It was collected in the Indian Creek, 35 site (Cacops Bonebed), from the Arroyo Formation of the Clear Fork Group, Baylor County of Texas, dating to the early Kungurian stage of the Cisuralian Epoch, about 279.5-272.5 million years ago. Many well preserved specimens from the same locality and horizon of the type specimen, including FMNH UR 2423, nearly complete skull and mandibles, MCZ 1926, complete skull and mandibles and FMNH P 12841, partial skeleton, are referred to V. brevirostris. One articulated skeleton with bite marks was found in southwest of Abilene (Arroyo Formation), Taylor County of Texas. Specimens (OMNH 73156-73178) of V. brevirostris were also collected in the Richards Spur, from the Garber Formation (Dolese Brothers Limestone Quarry) of the Sumner Group, Comanche County of Oklahoma, dating to the same age. Those remains came from at least three individuals, and represents the first varanodontine material from the Richards Spur. Finally, TMM 43628-1, a partial skeleton with nearly complete skull, was collected in the Mud Hill locality, from the Vale Formation of the Clear Fork Group, Taylor County, also dating to the same age. Varanops is the type genus of the famiy Varanopidae. Cladistic analysis performed by Nicolás E. Campione and Robert R. Reisz in 2010 suggests that Varanops is a derived varanodontine, sister taxon to the clade formed by Varanodon and Watongia.
Репродукции (1, 2, 3, 4, 5, 6):
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 07:42 pm [industrialterro] [Link] |
Secodontosaurus Секодонтозавр (Secodontosaurus) — необычный сфенакодонтный пеликозавр раннепермской эпохи. От других сфенакодонтов отличается длинной низкой мордой (напоминает морду рыбоядных крокодилов). Длина черепа около 30 см. Скорее всего, питался рыбой. Мог питаться роющими животными (вытаскивал их из нор?), на что указывает необычное для рыбоядного животного строение зубов и относительно сильные челюсти. 2 вида, из ранней перми Северной Америки (Техас). Типовой вид — S. obtusidens, описан Э.Д. Коупом в 1880 г как вид рода "диметродон", в особый род выделен А.Ш. Ромером в 1936 году.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 07:35 pm [industrialterro] [Link] |
Varanosaurus Varanosaurus ('monitor lizard') is an extinct genus of early pelycosaur synapsid that lived during the early Permian (260 million years ago). As its name implies, Varanosaurus looked very similar to present-day monitor lizards. It had a flattened, elongated skull and a pointed snout with a row of sharp teeth, including two pairs of conspicuous pseudocanines, implying that it was an active predator. It was a small, nimble reptile, up to 1.5 metres (4.9 ft) long. Varanosaurus probably lived in swamps, competing with the larger Ophiacodon for food.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 07:16 pm [industrialterro] [Link] |
Oedaleops Oedaleops is an extinct genus of the family Eothyrididae. It was closely related to Eothyris. Like Eothyris, it was probably an insectivore. Skull fragments from three individuals and some limb elements were found in the Abo Cutter formation in New Mexico. All remains belong to the single known species Oedaleops campi. It is dated to the Sakmarian age of the early Permian.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| October 30th, 2011 | |
| 02:34 pm [industrialterro] [Link] |
Eothyris Eothyris is an extinct genus of the family Eothyrididae. It was an insectivorous synapsid. It was closely related to Oedaleops. Only one species is known, Eothyris parkeyi (Romer), from a single skull, found in the Belle Plains Formation in Texas. It belongs to the Artinskian age of the Early Permian. Эотириды (Eothyrididae) или Эотиридиды — семейство крайне примитивных пеликозавров раннепермской эпохи. Близки к казеидам, могут быть их предками. Вместе с казеидами образуют группу Caseasauria. М. Ф. Ивахненко отрицает принадлежность казеид, эотирид, офиакодонтов и варанопсеид к синапсидной ветви амниот. Для этих животных им предложен подкласс Ophiacomorpha. Это связано с особенностями строения височной области этих групп, отличной от таковой настоящих синапсид — тероморфов. Эотириды отличаются короткой мордой, большими глазницами, череп низкий и широкий, височное окно небольшое, ноздри крупные. Присутствуют «рога» — небольшие выросты надвисочных костей. Зубы простые конические, есть пара «клыков» в каждой половине верхней челюсти. Зубной ряд прямой, на уровне нижнечелюстного сустава. Длина черепа не более 10 см. Скелет неизвестен, за исключением фрагментов конечностей. По всем признакам эотириды — самые примитивные пеликозавры. Но они появляются в палеонтологической летописи удивительно поздно. Известны два монотипических рода: Эотирис (Eothyris parkeyi) — описан А. Ш. Ромером в 1937 году из ранней перми (артинская эпоха) формации Бель Плэйнс Техаса. Эдалеопс (Oedaleops campi) — описан У. Лангстоном в 1965 году из формации Або Катлер (сакмарий) Нью Мексико. Вероятно, эотириды питались насекомыми и мелкими тетраподами, обитая вдали от водоемов на равнинах и водоразделах. Это объясняет их отсутствие в большинстве известных раннепермских и позднекарбоновых фаун.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 02:17 pm [industrialterro] [Link] |
Sphenacodon Сфенакодонты (Sphenacodontia) — семейство вымерших хищных синапсид подотряда эупеликозавров. Хищные формы, обычно крупного размера (до 4 −4,5 метров). Зубы сидят в глубоких ячейках, есть клыкоподобные зубы верхней челюсти, зубы пильчатые. Из-за мощных клыков верхнечелюстная кость разрастается высоко вверх, отделяет слезную кость от наружных ноздрей. Челюстной сустав находится ниже уровня зубного ряда. Есть угловой отросток нижней челюсти и направленный вниз ретроартикулярный отросток. Это представляет раннюю стадию эволюции среднего уха млекопитающих. Конечности стройные, вероятно, животные были весьма подвижны. Обычно выделяется единственное семейство — Sphenacodontidae. Хаптодусы иногда рассматриваются как группа более примитивных форм вне данного семейства. Собственно сфенакодонты отличаются высокими остистыми отростками туловищных позвонков, образующих у ряда форм высокий «парус». Наиболее известны роды диметродон и сфенакодон из ранней перми Северной Америки и Западной Европы. Сфенакодонты интересны как предки терапсид. Конкретные промежуточные формы между ними не найдены, но все анатомические особенности указывают на существование таких форм. Возможно, они обитали в тех областях, где захоронение ископаемых остатков было практически невозможным (например, вдали от водоемов, на водоразделах), в то время как почти все пеликозавры населяли зону влажного тропического климата и берега водоемов. Сфенакодон (Sphenacodon) — крупный хищный пеликозавр раннепермской эпохи. Внешне напоминал широко известного диметродона, но не имел столь развитого спинного «паруса». Тем не менее, остистые отростки довольно длинные. Типовой вид — S. ferox — описан О. Ч. Маршем в 1878 г. В 1937 году А. Ромер описал второй вид — S. ferocior. Оба вида сфенакодонов найдены в позднекарбоновых - раннепермских отложениях (формация Або/Катлер, около 285—288 млн л.н.) Нью-Мексико. Sphenacodon ferox достигал 177 см в длину и обладал низкими остистыми отростками. Второй вид, Sphenacodon ferocior был гораздо крупнее (до 2,25 м длиной) и массивнее. У него есть невысокий «парус». Второй вид отмечен для более высоких горизонтов данной формации. Несомненно, сфенакодон был главным хищником своего времени и места. Он обитал в болотистых лесах и мог питаться другими наземными позвоночными. Интересно, что в слоях Або/Катлер найден лишь один мелкий вид диметродона — он не мог конкурировать со сфенакодоном. Возможно, местообитание сфенакодонов было каким-то образом изолировано от других территорий Северной Америки, где доминирующим хищником был диметродон. К роду сфенакодон относят также описанного Ф. фон Хюне в 1908 году пеликозавра из раннепермских отложений Англии. Первоначально его выделяли в род Oxyodon, но сейчас этот вид называется Sphenacodon britannicus.
Репродукции (1, 2, 3, 4, 5, 6):
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 02:02 pm [industrialterro] [Link] |
Ctenorhachis Ctenorhachis (Greek for "comb spine") is an extinct genus of the family Sphenacodontidae. Ctenorhachis was related to Dimetrodon, but did not belong to the same subfamily as Dimetrodon and Sphenacodon, being a more basal member of Sphenacodontidae. Ctenorhachis lived in the Early Permian epoch. Two specimens are known that have been found from the Wichita Group outcropping in Baylor and Archer counties, north-central Texas. Only the vertebrae and pelvis are known. Articulated vertebrae from the holotype specimen possess blade like neural spines that are greatly enlarged, although not nearly to the extent that can be seen in more derived sphenacodontds such as Dimetrodon and Secodontosaurus, in which they form a large sail. The pelvis is nearly identical to that of Dimetrodon. As suggested in the original description of the genus, Ctenorhachis may represent a short-spined sexual dimorph, although the authors find this unlikely.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 01:36 pm [industrialterro] [Link] |
Bathygnathus Батигнат (Bathygnathus borealis) — сфенакодонтный пеликозавр из ранней перми Полярной Канады (остров Принца Эдварда). Описан Джозефом Лейди в 1854 году на основании куска челюсти из красных песчаников этого острова. Лейди решил, что фрагмент кости с зубами является дистальным концом правой ветви нижней челюсти крупного хищного динозавра. Он назвал его «Батигнат» — «тяжелая челюсть», поскольку высота челюсти была велика для ее длины. Согласно описанию Лейди, в челюсти сохранились семь крупных зубов неравной величины, высота фрагмента больше 10 см (4 дюйма), длина — более 18 см (7,25 дюйма); задняя часть фрагмента чрезвычайно тонка, что указывает на существовавший контакт с другими костями (с надугловой костью, по мнению автора). Наружная поверхность челюсти покрыта тонкой структурой из мелких бороздок. Зубы уплощенные, напоминают зубы современных варанов. Лейди отметил, что зубы могли замещаться изнутри кнаружи. На основании своего описания Лейди заключил, что возраст красного песчаника острова Принц Эдвард — триасовый. Прочие геологи не были вполне согласны с ним, но наличие остатков динозавра не оставляло возможности другого толкования, тем более, что Э. Д. Коуп был согласен с определением Лейди. В 1905 году Э. Кейз и Ф. фон Хюне одновременно пришли к заключению, что челюсть батигната — левая верхнечелюстная кость (максилла) крупного хищного пеликозавра, сходного с диметродоном. Соответственно, возраст красных песчаников Полярной Канады в этом районе оказался раннепермским, что подтвердили и найденные остатки растений. Судя по размерам кости, длина всего черепа могла быть до 45-50 см, что сравнимо с крупнейшими видами диметродонов. Следует отметить, что по строению черепа крупные сфенакодонты (диметродон, сфенакодон, ктеноспондил) практически неотличимы. Поэтому истинное систематическое положение батигната внутри семейства и его облик (наличие «паруса») остается неизвестным. Так или иначе, батигнат показывает, что распространение сфенакодонтов в начале пермского периода было очень широким.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 12:46 pm [industrialterro] [Link] |
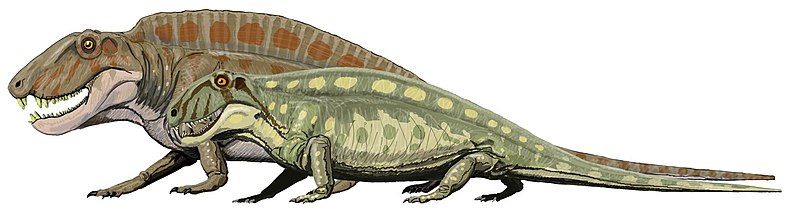
Dimetrodon Диметродон (Dimetrodon) — род вымерших хищных синапсид из семейства сфенакодонтов, живших в пермский период 280—265 млн лет назад. Эволюционно находится ближе к млекопитающим, чем к истинным рептилиям. Достигал в длину 2,7—4,0 метров и был крупнейшим хищником своего времени. В отличие от рептилий имел зубы двух типов (резцы и клыки), однако не подобранные под туловище ноги. Наиболее яркой чертой диметродона является парус из натянутой на выросты спинных позвонков кожи. Подобные паруса развились у различных доисторических животных (амфибии Platyhystrix, пеликозавра Edaphosaurus, динозавра Spinosaurus) и служили терморегуляторами. Подсчитано, что 200-килограммовый хладнокровный диметродон разогревался бы без паруса с 26 °C до 32 °C за 205 минут, а с парусом за 80 минут. По другим версиям парус использовался в брачных играх, служил камуфляжем среди вертикальных стеблей растений или настоящим парусом при плавании. «Парус» развивался постепенно в течение жизни, известны находки молодых особей диметродона с невысокими остистыми отростками. Молодые особи, по-видимому, обитали в зарослях по берегам водоемов, взрослые — жили в лесах и на равнинах. Тем не менее, бытующее в популярных книгах изображение диметродона на фоне пустыни представляется нереальным — не будучи настоящей рептилией, он не мог бы выжить в жаркой сухой местности. Диметродоны питались любыми животными, которых могли одолеть. Интересно, что в условиях примерно равного соотношения биомассы хищников и растительноядных (характерного для раннепермских фаун), основу питания в отдельные периоды могла составлять рыба. В 2008 году команда Р. Бэккера обнаружила четкие признаки того, что диметродоны поедали пресноводных акул — ксенакантид. Возможно, акулы оставались в пересыхающих прудах, где становились жертвой пеликозавров. Описаны 12−13 видов диметродона: Хорошо известны:
Описаны по фрагментам:
Слишком фрагментарны: D. kempae (Romer 1937) — Кунгурский ярус. В последние годы фрагменты позвонков, предположительно диметродонов обнаружены в пермских отложениях Южной Америки.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13):
Размеры в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9, 10):
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 12:36 pm [industrialterro] [Link] |
Mycterosaurus Mycterosaurus was a pelycosaur belonging to the family Varanopidae of the subfamily Mycterosaurinae. Mycterosaurus is the most primitive member of its family. It lacks some features that its advanced relatives have. Варанопсеиды (Varanopseidae) или варанопиды (Varanopidae) – семейство мелких хищных пеликозавров позднего карбона – средней перми. Последние из пеликозавров и единственная группа примитивных синапсид, обнаруженная в Южной Африке. В целом напоминали крупных ящериц типа современных варанов (от ящериц их отличало несколько более массивное телосложение). Зубы острые, многочисленные, сжатые с боков. Морда обычно длинная, глаза большие. У варанодона описано предглазничное окно (как у архозавров). Обитали на равнинах, питались мелкими животными, мелкие виды – насекомоядные. У рода эллиотсмития (Elliotsmithia) имелись мелкие костные чешуи на спине. Миктерозавр (Mycterosaurus) – ранняя пермь Северной Америки (Техас). Размеры варанопид составляли от 30 - 50 см (археовенатор, пиозия) до 1,5 метров (варанодон, варанопс, рутиромия). В 1974 году Э.Олсон описал из «средней» перми свиты Чикаша в Оклахоме разрозненные останки синапсиды, названной «ватонгия» (Watongia). Олсон считал ватонгию примитивным горгонопсом. Сейчас доказано, что ватонгия – скорее всего, гигантский варанопид (длиной не менее 2 метров). Вероятно, в конце своей истории в Северной Америке варанопиды заняли положение доминирующего хищника. Ватонгия могла охотиться на крупную добычу (например, на молодь казеидных пеликозавров, найденных в тех же отложениях). Крупные поздние варанопиды - варанопс, варанодон и ватонгия формируют четкую естественную группу. Последние варанопиды – эллиотсмитии – были мелкими насекомоядными животными. Недавно из поздней перми Южной Африки описан скелет взрослого варанопсеида, окруженный скелетами меньших по размеру особей (детенышей?). По-видимому, вся "компания" погибла в норе во время наводнения. К варанопидам принадлежит также гелеозавр (Heleosaurus). Это ящерицеобразное животное из поздней перми (зона Tropidostoma) Южной Африки долгое время считалось примитивным архозавром (именно так гелеозавра изображал известный палеохудожник Дж. Сиббик). Сейчас установлено, что гелеозавр - один из поздних панцирных варанопид, сходный с эллиотсмитией. Более того, это может быть то же самое животное. В таком случае название Heleosaurus имеет приоритет. В связи с этим "семью" варанопсеид из Южной Африки сейчас относят к этому роду. М.Ф. Ивахненко отрицает родство варанопсеид со сфенакодонтами, сближая их с офиакодонтами и казеозаврами в составе подкласса Ophiacomorpha (офиакоморфы).
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 12:19 pm [industrialterro] [Link] |
Ennatosaurus Ennatosaurus (meaning "the 9th reptile") was a mammal-like reptile that lived in Russia during the Wordian stage of the Permian period. It is included in the synapsid clade Caseidae in the order Pelycosauria. Ennatosaurus was herbivorous, possibly using its broad forefeet to dig at plants. Like all caseids, Ennatosaurus had a small head compared to its wide, lizard-like body. Its mouth was lined with blunt, peg-like teeth. It lived alongside other Permian creatures, such as the herbivorous Nyctiphruretus and the carnivorous Biarmosuchus. The adult size for Ennatosaurus is unknown. Fossil remains show an animal about the size of a cat, but these are likely juvenile specimens, the adults growing to sizes that may have rivaled those of 20-foot (6.1 m)-long cousin, Cotylorhynchus. Ennatosaurus is known from only one fossil site, where several juveniles were buried simultaneously in sand. One adult skull was found among the juvenile skeletons. Эннатозавр (Ennatosaurus) — «средняя» пермь Архангельской области, Россия (мезенская фауна, соответствует по возрасту Очерской фауне). Близок к американским родам типа котилоринха. Длина черепа до 20 см (больше, чем у котилоринха), но, судя по посткраниальным остаткам, обладал довольно крупной головой, поэтому общую длину оценивают в 1,5 метра. Не исключено, что полные реконструкции основаны на соединении черепа взрослой особи и скелета молодого животного. Вероятно, последний из казеид.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 11:30 am [industrialterro] [Link] |
Cotylorhynchus Cotylorhynchus was a very large mammal-like reptile (more accurately synapsid) that lived in the southern part of what is now North America during the Early Permian period and persisted until the late-Mid Permian (about 265 mya). Cotylorhynchus are the most well known of the synapsid clade Caseidae and are in the order Pelycosauria. The Cotylorhynchus were the largest pelycosaurs, the largest terrestrial vertebrates of their time, as well as the terrestrial vertebrates up to that time. They were herbivores, and because of their enormous size they probably did not fear any of the carnivores. Cotylorhynchus was a massively built animal with a disproportionately small head and a huge barrel-shaped body. They could grow to lengths of up to 20 feet and could achieve weights of up to 2 tons. Their skeletal features included a massive scapulocoracoid, humeri with large flared ends, stout forearm bones and broad, robust hands that had large claws. Certain features of their hands indicate that they had to dig considerably to obtain their food supply and also they may have used these features to dig burrows for shelter or safety. Their digits were believed to have a considerable range of motion and large retractor processes on the ventral surfaces of the unguals allowed them to flex their claws with powerful motions. Also, the articulatory surfaces of their phalanges were oblique to the bone's long axis rather than perpendicular to it. This allowed for much more surface area for the flexor muscles. Their skulls are distinctive in the presence of large temporal openings and very large nostril openings, which could have been utilized for better breathing or may have housed some sort of sensory or moisture conserving organ. Also they featured large pineal openings and a snout or upper jaw that overhangs the row of teeth to form a projecting rostrum. Rounded deep pits and possibly large depressions were present on the outer surface of the skull. Their teeth were very similar to those of iguanas with posterior marginal teeth that bore a longitudinal row of cusps. Котилоринх (Cotylorhynchus) — верхние горизонты ранней перми (кунгурская эпоха) Техас и Оклахома. Крупнейший представитель казеид. Типовой вид (Cotylorhyncus romeri) мог достигать почти 3 метров в длину. Самый крупный вид — Cotylorhynchus hancocki, до 6 метров в длину и весом до 2 тонн. Никогда ранее растительноядное наземное позвоночное не достигало таких размеров. Вид был описан Олсоном и Бирбауэром в 1953 году из среднепермских слоев свиты Сан-Анжело (кунгурская эпоха, около 270 млн л.н.) в Техасе. Типовой вид котилоринха описан по хорошо сохранившимся полным скелетам. Это неуклюжее животное с очень маленькой головой, уплощенным бочкообразным телом, длинным хвостом и очень мощными короткими ногами. Очень мощные когти на передних и задних лапах, фаланговая формула сокращена. Котилоринх Хэнкока плохо известен, но отличался более мощными конечностями, когти передних лап достигали 7—8 см в длину. Третий вид — Cotylorhyncus bransoni — из средней части формации Флауэрпот в Оклахоме. Он близок по размеру к типовому виду, но отличался рядом деталей строения, в том числе более короткими конечностями.
Репродукции (1, 2, 3, 4, 5, 6, 7):
Размеры в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 11:11 am [industrialterro] [Link] |
Casea Casea is an extinct genus of pelycosaur synapsid which was about 1.2 metres (4 ft) long, slightly smaller than the otherwise very similar Caseoides. Casea was one of the first terrestrial herbivores, sharing its world with animals such as Dimetrodon and Eryops. Casea had a heavy, rotund, body and a small skull. Its rib cage was greatly expanded, presumably to make space for a large, plant-fermening gut. Like other caseids, it lacked teeth in its lower jaw, and had blunt teeth in the upper jaw. These adaptations indicate that Casea was a herbivore, feeding on relatively tough plants, such as ferns. Казея (Casea) — ранняя пермь Техаса и Франции. Типовой вид (C. broilii) был описан С. У. Виллистоном в 1910 году из раннепермской формации Арройо в Техасе. Типовой вид — мелкое животное, длиной до 1,5 метров. Череп небольшой, конечности относительно короткие, тело бочкообразное. Второй вид — C. nicholsi происходит из более поздних отложений формации Вале в Техасе. Крупнее типового вида, с более мощными конечностями. Череп плохо известен. Третий американский вид — Casea halselli, в дину мог достигать 2-3 метра. Происходит из еще более поздней формации Чоза в Техасе. В 1974 году из раннепермских отложений Франции был описан еще один вид — C. rutena, мелкое животное, близкое к типовому виду рода.
Размеры в сравнении с человеком:
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| 11:00 am [industrialterro] [Link] |
Angelosaurus Angelosaurus dolani ('Dolan's Angel lizard', Olson & Berrbower 1953) was a pelycosaur (an extinct clade) of reptile. It is known from a partial skull from the Middle San Angelo Formation, Knox County, Texas, USA. It dates from the Kungurian (Early Permian). It was from three to three and a half metres long and weighed about 300 kilograms. It belongs to the family Caseidae. Like its contemporary, another caseid, Cotylorhynchus, it was large, heavily built, and herbivorous. It also shared its environment with the caseids Caseoides and Caseopsis. It may have been preyed upon by the large sphenacodont pelycosaur Dimetrodon. Like all caseids, Angelosaurus was an evolutionarily conservative reptile which would appear to have been one of the dominant herbivores of the time, occupying a similar ecological role to cows. Caseid skulls are distinguished from other pelycosaurs by large temporal openings, large nasal and pineal openings and an upper jaw that distinctively overhangs the teeth of the lower jaw.The skull is pitted. The teeth are uniform. The body was probably barrel-shaped, like other caseids, with massive limbs and small cervical vertebrae. And unlike the more well-known pelycosaurs such as Dimetrodon and Edaphosaurus, caseids do not have the exaggerated elongation of the vertebrae that form a sail. Two other species of Angelosaurus have been described so far: Angelosaurus greeni Olson 1962, and Angelosaurus romeri Olson and Barghusen, 1962. Казеиды (Caseidae) — растительноядные пеликозавры, существовавшие в течение всего пермского периода. Появились в конце раннепермской эпохи, дожили до середины перми. «Паруса» нет, тело массивное, конечности обычно короткие и толстые, хвост относительно длинный. Голова небольшая, у поздних крупных видов — очень маленькая. Зубы однообразные, выделяются лишь крупные конусовидные резцы; коронки лопатообразные зазубренные (как у парейазавров). Окклюзии зубов нет, животное могло только отрывать куски пищи и проглатывать ее. Развиты острые многочисленные небные зубы. Крупное височное отверстие черепа, огромные ноздри (ноздря была в значительной степени перекрыта септомаксиллой, но она редко сохраняется). Крупное пинеальное отверстие («теменной глаз»). Череп скульптирован мелкими ямками. Возле ноздри могла находиться какая-то (возможно, солевая) железа. М. Ф. Ивахненко включает казеид вместе с офиакодонтами и варанопидами в состав особого подкласса Ophiacomorpha, не имеющего прямого родства с другими «пеликозаврами». Ангелозавр (Angelosaurus) — происходит из верхних горизонтов ранней перми (формации Флауэрпот и Сан-Анжело, кунгурская эпоха) Оклахомы и Техаса. Характеризуется чрезвычайно массивным скелетом, особенно массивны проксимальные элементы конечностей. Три вида, крупнейший (A. greeni) достигал почти 3 метров в длину. Известны преимущественно по остаткам посткраниального скелета и редким фрагментам черепа. Интересны очень тупые когти, в отличие, например, от котилоринха.
Tags: Вымершие синапсиды, Пермь, пеликозавры |
| October 8th, 2011 | |
| 07:38 pm [industrialterro] [Link] |
Ophiacodon Офиакодон (Ophiacodon) – примитивный хищный пеликозавр из позднего карбона – ранней перми Северной Америки и, возможно, Англии. Относится к группе офиакодонтов (семейство Ophiacodontidae). Самый крупный и самый известный представитель группы. Череп очень крупный, длинный и высокий, множество острых зубов. «Клыки» мало выделяются среди прочих зубов. Тело у крупных видов массивное, ноги короткие, слабо окостеневшие, иногда с тупыми когтями. Около 6 видов, крупнейшие из которых (O. retroversus) достигали в длину 2,5 – 3,6 метра. Интересная особенность – наличие у некоторых крупных особей двух височных отверстий, описан также экземпляр с двумя отверстиями только на одной стороне (с другой было одно отверстие). Считается рыбоядным, но у крупных особей слишком высокий череп. Вероятно, был полуводным хищником, питавшимся самой разнообразной добычей (например, земноводными или дохлой рыбой). Описан О.Ч. Маршем в 1878 году из ранней перми Техаса. Множество синонимов рода упоминается в старой литературе (Diopaeus, Theropleura, Therosaurus, Poliosaurus и др.). Ophiacodon mirus Marsh, 1878 — типовой вид, описан из нижней перми (сакмарский ярус) формации Або/Катлер, Нью-Мексико. Известен по неполному скелету. Некрупный вид, общая длина тела около 1,6 м. Из нижней перми (предположительно, отенский ярус) Англии описан пеликозавр, отнесенный к Ophiacodon sp. Он известен по плохо сохранившемуся фрагменту левой зубной кости. Изначально Мурчисон и Стрикленд (1840) приняли этот образец за верхнечелюстную кость рыбы. Челюсть тонкая и очень низкая, постепенно расширяющаяся спереди назад. Сохранен симфизный отдел и полный зубной ряд, но большая часть вентральной части отсутствует. Длина фрагмента составляет 122 мм. В зубном ряду размещалось приблизительно 34 зуба. Первый зуб в ряду, вероятно, был большим, как второй и третий. Третий зуб полностью сохранился, его длина состаляет 11 мм. Четвертая альвеола, вероятно, также вмещала большой зуб. Зубы с 5 по 26 примерно одинаковы по размеру, длиной 5 мм. Зубы с 27 по 34 намного меньше, уменьшаются в размере спереди назад; длина наименьшего зуба — 2 мм. Зубы не сжаты с боков и не имеют режущих краев; зубы с 5 по 34 конические и колышковидные, с тупыми вершинами. Полностью сохранившийся увеличенный третий зуб немного загнут и заострен. Структура челюсти и строение зубов европейского Ophiacodon sp. типичны для офиакодонтов и отличаются от таковых у сфенакодонтов. Группа увеличенных зубов на переднем конце нижней челюсти характерна для пеликозавров. Зубы мельче и более многочисленны, чем у сфенакодонтов, а в зубном ряду есть много промежутков. Это также особенность, присущая офиакодонтам, и, вероятно, связанная с медленной сменой зубов. По структуре челюсть напоминает нижнюю челюсть пеликозавров из рода Ophiacodon. До обнаружения этой находки представители рода, даже наиболее примитивные формы из каменноугольного периода, были известны только в Северной Америке. Длина черепа европейского офиакодона могла равняться приблизительно 185 мм, что значительно меньше, чем у любой из известных американских форм. Однако, возможно, остатки принадлежат молодому животному. Количество зубов также меньше, чем у американских видов, хотя и превышает число, характерное для сфенакодонтов. Кроме меньшего размера и числа зубов, в остальном челюсть наиболее сходна с челюстью Ophiacodon uniformis, формы из отенского яруса формации Уичита. Малое число отличительных признаков не позволяет отнести европейскую форму к отдельному или одному из описанных видов рода Ophiacodon.
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие синапсиды, Карбон, пеликозавры |
| 07:29 pm [industrialterro] [Link] |
Ianthasaurus Ianthasaurus was a small edaphosaurid from the Late Pennsylvanian. Ianthasaurus lacks many of the spectacular specializations seen in Edaphosaurus. For example, the marginal dentition of Ianthasaurus is similar to that of insectivorous reptiles, with slender conical teeth which are slightly recurved at the tips, and there is a slight development of a caniniform region. The palatal and mandibular dentition is unspecialized, and there are no batteries of teeth for crushing of plant materials. Also unlike Edaphosaurus, Ianthasaurus was lightly built and was probably quite agile. The skull was similar to that to Haptodus, a sphenacodontid, though they were distantly related. Самый примитивный из эдафозавров — янтазавр (Ianthasaurus), некрупное животное из позднекаменноугольных отложений Северной Америки и Европы. У него сохраняются длинные зубы, указывающие на возможное питание насекомыми. Длина около 1 метра, «парус» низкий. Типовой вид - Ianthasaurus hardesti из позднего карбона Гарнетта в Канзасе. Вид описан Рейсом и Берманом в 1985 году. Изучение остатков мелких эдафозавров из позднего карбона-ранней перми Западной Европы (Чехия и Германия) показало их возможную принадлежность к данному роду. Это I. mirabilis, описанный Фритчем в 1895 году и I. credneri, описанный Йекелем в 1910 году. Впрочем, посткраниальные остатки янтазавра практически неотличимы от таковых мелких эдафозавров.
Tags: Вымершие синапсиды, Карбон, пеликозавры |
| October 6th, 2011 | |
| 11:42 pm [industrialterro] [Link] |
Haptodus Haptodus was a small sphenacodont, a lineage that includes therapsids. It was at least 1.5 metres (5 ft) in length. It lived from Latest Carboniferous to Early Permian, in the equatorial Pangea. It was a medium-sized predator, feeding on insects and small vertebrates. It is one of the most primitive sphenacodontid and it is related to other sphenacodontids (or it may have been ancestral to them), such as Dimetrodon. It shares many structural features of the skull and skeleton with the more specialised sphenacodontids. Unlike Dimetrodon, it lacked the "sail". Хаптодусы (Haptodontinae) — подсемейство хищных пеликозавров семейства Sphenacodontidae. Наиболее примитивные из сфенакодонтов. Известны из позднего карбона — ранней перми Северной Америки и Европы. Животные средних размеров (длиной до 1,5 −2 метров). Череп высокий, короткий, зубы мощные. «Паруса» нет. По образу жизни напоминали современных хищных ящериц, обитали в тропических лесах. Обычно выделяют лишь один род хаптодус (Haptodus). Тем не менее, иногда род разделяют на 4 рода (подрода): Катлерия — Cutleria (с единственным видом C. wilmarthi) — из ранней перми (формация Або/Катлер, сакмарский век) Нью-Мексико. Хаптодус — Haptodus (H. baylei, H. grandis из позднего карбона — ранней перми Западной Европы, длиной до 2 метров). Сюда относится также самый примитивный и мелкий вид — H. garnettensis из позднего карбона (касимовский век) Канзаса. Длина до 60 см. Палеогаттерия — Palaeohatteria longicaudata — ранняя пермь (сакмарский век) Германии. Один из наиболее известных видов. Описан Креднером в 1888 году. Изначально считался диапсидной рептилией, предком современной гаттерии. Длина до 0,6—1 м. Большинство остатков принадлежат молодым особям. Пантелозавр — Pantelosaurus saxonicus — ранняя пермь (ассельский век) Германии. Известен по скоплению 6 скелетов молодых особей, найденному в шахте близ Фрейталя, возле города Дрезден. Массивное высокоголовое животное, длина взрослых доходила до 1,5 метров. Возможно, является синонимом вида Haptodus baylei. Хаптодусы могут быть исходной группой для происхождения не только настоящих сфенакодонтов (таких, как диметродон), но и терапсид. Не исключено, что загадочный пеликозавр тетрацератопс близок к хаптодусам.
Репродукции:
Собственно сам хаптодус (1, 2, 3, 4, 5):
Tags: Вымершие синапсиды, Карбон, пеликозавры |
| 11:24 pm [industrialterro] [Link] |
Edaphosaurus Edaphosaurus (pronounced /ˌɛdəfɵˈsɔrəs/) is a genus of prehistoric synapsid which lived around 303 to 265 million years ago, during the late Carboniferous to early Permian periods. The name Edaphosaurus means "ground lizard" and is derived from the Greek edaphos/εδαφος ("ground, land") and σαυρος/sauros ("lizard"). This genus was first known from remains discovered in North America, in the late 19th century, and described by American paleontologist Edward Drinker Cope. More recently other fossils attributed to Edaphosaurus have been found in the Czech Republic and Slovakia. The most characteristic feature of Edaphosaurus is a sail on its back similar to that of other synapsids of the same time, such as the large apex predator Dimetrodon. However, the sail on Edaphosaurus is different in shape and morphology. Along with the reptile-like family Diadectidae, Edaphosaurus is one of the earliest known large plant-eating tetrapods (land-living vertebrates). Early descriptions, referring to its sharp teeth, suggest that the synapsid may have fed on small invertebrates, such as mollusks, although paleontologists now point out that Edaphosaurus also shows herbivore characteristics. The head of Edaphosaurus was short and shallow, and remarkably small compared to its body size. The deep lower jaw had powerful muscles and the frontal teeth were barbed at the cutting edges, helping Edaphosaurus to cut bite-sized pieces from tough terrestrial plants. The palate and inside the lower jaw are covered with small teeth forming a large biting surface. The sail along the back of Edaphosaurus was supported by hugely elongated neural spines from neck to lumbar region. As compared with the sail of Dimetrodon, the vertebral spines are shorter and heavier and bear numerous small cross bars. Edaphosaurus species measured from 1 m (3 ft) to almost 3.5 m (11 ft) in length and weighed over 300 kilograms (660 lb). This large herbivore had a massive wide body, thick tail and short limbs which show that was a slow moving animal. Эдафозавры (Edaphosauridae) — семейство растительноядных пеликозавров. Обитали в Северной Америке и Европе в конце каменноугольного — начале пермского периодов. Одни из первых крупных доминирующих растительноядных (могли питаться отмершей растительностью). Голова маленькая, зубы колышковидные, равного размера. Небо и внутренняя сторона нижней челюсти покрыты дополнительными зубами. Есть глубокая нижняя вырезка скуловой дуги. Возможно, имелись мягкие щеки, позволявшие удерживать пищу во рту. Конечности короткие, тело массивное, широкая грудная клетка. У всех эдафозавров был «парус», образованный остистыми отростками спинных позвонков. На отростках имелись поперечные выросты, верхушки нескольких передних отростков часто расширены. Вероятно, как и у сфенакодонтов, «парус» служил для терморегуляции, поперечные отростки могли увеличивать его площадь. Кости "паруса" были полыми. Нельзя также исключить, что у основания "паруса" могли откладываться запасы жира. Собственно эдафозавр (Edaphosaurus) — крупное растительноядное, длина некоторых видов доходила до 3,5 метров. 6 −9 видов из позднего карбона — ранней перми Северной Америки. Наиболее крупные виды — E. cruciger и E. pogonias из раннепермских отложений Техаса, близок по размеру более примитивный E. boanerges. Эти виды были современниками диметродонов и могли служить их добычей. Род был описан Э.Д. Коупом в 1882 году по деформированному черепу из ранней перми Техаса. Он считался близким к "котилозаврам", к которым тогда относили всех примитивных рептилий. В 1886 году Коуп описал под названием "наозавр" (Naosaurus) "парусного" пеликозавра из тех же отложений. Считалось, что череп наозавра не отличался от черепа диметродона, но "парус" нес поперечные отростки. Такая реконструкция "наозавра" часто встречается в старой литературе (вплоть до 1930-х годов!). В 1907 году Э.Кейз предположил, на основании новых находок, что череп "эдафозавра" принадлежит "наозавру". Впоследствии это блестяще подтвердилось. Таким образом, "наозавр" был результатом искусственного соединения черепа диметродона с черепом эдафозавра. Эдафозавр, также известный как Edaphosaurus, является растительноядным представителем семейства пеликозавров. Особи некоторых видов этого семейства достигали в длину от 1 до 3,5 метров. Наиболее яркая отличительная черта – парус на спине, подобный наличествующим у других синапсидов того времени. Эдафозавры являются одними из первых известных науке больших травоядных четвероногих; вместе с другими синапсидами они приходятся далекими предками современным млекопитающим. Название вида в переводе с греческого значит «земляная ящерица». Приземистый силуэт, массивная туша(крупные особи весили более 300 кг) с широкой грудной клетью, короткие лапы, вынесенные в стороны от туловища, длина хвоста чуть менее длины тела – внешний вид эдафозавра был более или менее типичен для пеликозавров. Парус, украшавший спину животного, использовался, по мнению ученых, для терморегуляции (также возможно применение паруса в целях внутривидовой демонстрации для устрашения самок и привлечения самцов в период спаривания). Парус был образован костными отростками спинных позвонков, причем кости паруса были полыми. Возможно, у основания паруса скапливались запасы жира. Любопытно, что у этого вида на отростках, составляющих «основу» паруса, есть поперечные отростки для увеличения площади паруса, что, в свою очередь, приводит к улучшению терморегуляции. У ранних видов эдафозавров были острые и довольно длинные зубы. Это говорит о том, что их предки могли питаться небольшими беспозвоночными вроде моллюсков, однако вполсдествии перешли к вегетарианству. Голова у типичных эдафозавров была маленькая, в пасти размещались колышковидные зубы одинакового размера. Внутренняя сторона нижней челюсти и небо также были оснащены зубами. Предположительно, у этого вида могли быть мягкие щеки, благодаря чему животные удерживали во рту большее количество пищи. Что касается взаимодействия с другими видами того времени, эдафозавры служили пищей другому виду пеликозаврам – диметродонам. Что характерно, последние тоже обладали костным парусом, однако были стопроцентными и вполне успешными хищниками. Сейчас ученые располагают сведениями о том, что эдафозавры существовали 303-265 млн лет назад преимущественно в Северной Америке, несколько видов известно также из Европы(Чешская Республика и Словакия). Причиной вымирания этого вида стала, по-видимому, смена климата. Им на смену пришел другой вид растительноядных пеликозавров – казеиды, которые доминировали в этой нише на протяжении первой половины пермского периода.
Репродукции (1, 2, 3, 4, 5, 6, 7):
Ископаемые останки (1, 2, 3, 4, 5, 6, 7):
Tags: Вымершие синапсиды, Карбон, пеликозавры |













































/015_Dimetrodon.jpg)













.JPG)


















/Cotylo_01.jpg)


















































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}