[Recent Entries][Archive][Friends][User Info]
March 27th, 2012
| March 27th, 2012 | |
|---|---|
| 06:17 pm [industrialterro] [Link] |
Unaysaurus Unaysaurus — вымерший род динозавров принадлежащих к группе Sauropodomorpha, живших в позднем триасе около 225 миллионов назад (карнийский—норийский ярусы) на юге нынешней Бразилии. Типовой вид - Unaysaurus tolentinoi. Ископаемые останки, голотип UFSM 11069, были найдены в 1998 году в окрестностях города Санта-Мария. Находка была сделана Толентино Марафигой случайно, во время прогулки (видовое название дано в его честь). Окаменелости залегали в слоях формации Caturrita, относящейся к карнийскому—норийскому ярусам, и они состоят из почти полного черепа и некоторых фрагментов туловища и конечностей. В 2004 году вид был описан в научном журнале Zootaxa Александром Кельнером. Название рода происходит от слова unay - «черная вода», на языке тупи, ссылаясь на регион Agua Negra, на территории которого была сделана находка. Unaysaurus был относительно небольшим травоядным динозавром, достигавшим в длину 2,5 метров и весившим 75 кг. Внешне напоминал прозауроподов: он передвигался на двух ногах (также не исключено, что он мог опираться и на 4 конечности) и имел длинную шею. В современных классификациях динозавра относят к семейству Plateosauridae, но вляется ли он вдействительности членом группы Prosauropoda в настоящее время остается неясным. Бразильские ученые обнаружили хорошо сохранившиеся окаменелые останки динозавра неизвестного ранее вида, который обитал на территории южной Бразилии 225 миллионов лет назад. Как передает Reuters, высота этого травоядного ящера, который передвигался на задних лапах, составляла около 70 сантиметров, а длина - 2,5 метра.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, платеозавриды, прозавроподы, ящеротазовые |
| Time | Event |
| 06:48 pm [industrialterro] [Link] |
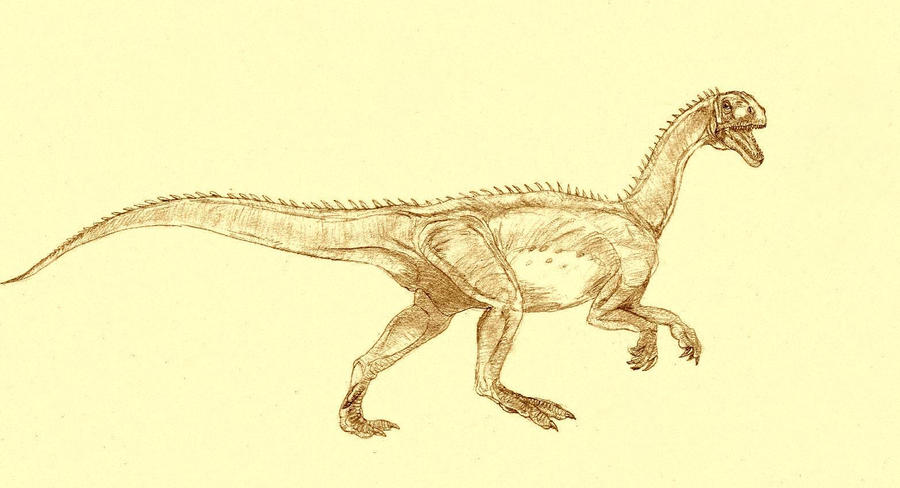
Zupaysaurus Зупайзавр (Zupaysaurus) — род вымерших ящеротазовых динозавров из подотряда тероподов, живших в позднем триасовом и раннем юрском периодах (около 216,5—203,6 миллионов лет назад), на территории нынешней Южной Америки. Окаменелости теропода были найдены в провинции Ла-Риоха (формация Los Colorados) в Аргентине. Впервые описан палеонтологами Аркуччи (Arcucci) и Кориа в 2003 году. Представлен одним видом — Z. rougieri. Хотя полный скелет до сих пор не был обнаружен, ученые считают Zupaysaurus двуногим хищником, достигавшим до 4 метров в длину. Также ученые предполагают возможное наличие двух параллельных гребней на его длиной морде. Zupaysaurus был динозавром среднего размера. Череп взрослой особи достигал 45 сантиметров в длину, что дает возможность ученым предположить общую длину животного, которая оценивается примерно в 4 метра от морды до кончика хвоста. Как и все известные тероподы, Zupaysaurus вероятно ходил только на задних конечностях, оставляя передние свободными для хватания добычи. Имел небольшой зазор, отделявший зубы предверхнечелюстных и верхнечелюстных костей верхней челюсти, таранная кость и кости пяточной лодыжки были слиты воедино, что отмечается также и у ранних тероподов. В верхней части черепа вероятно имел два тонких параллельных гребня, подобно другим тероподам, таким как Dilophosaurus и Megapnosaurus (Syntarsus) kayentakatae. Эти гребни предположительно были сформированы исключительно носовыми костями, в отличие от гребней многих других тероподов, у которых подобные образования дополнялись также и слезными костями. Ученые предполагают, что гребни, широко распространенные на черепах тероподов, возможно использовались ими в коммуникативных целях, таких как гендерное, групповое или видовое распознавание. Однако, более свежий анализ черепа подверг сомнению вероятность присутствия этих гребней в Zupaysaurus. Неопубликованное резюме, представленное на конференции, указывало на то, что структуры, первоначально идентифицированные как гребни, на самом деле, были фактически слезными костями, смещенными вверх во время процесса окаменения.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| Time | Event |
| 07:41 pm [industrialterro] [Link] |
Protoavis Протоавис (Protoavis) — название рода, предложенное для окаменелостей позднего триаса (225—210 млн лет), обнаруженных на северо-западе Техаса (США). В 1991 году протоавис был описан палеонтологом Шанкром Чаттерджи (Sankar Chatterjee) как примитивная птица, однако эта интерпретация была подвергнута сомнению. Если интерпретация Чаттерджи верна, то протоавис является древнейшей из известных птиц, на 60-75 млн лет старше археоптерикса. Выделяется единственный вид Protoavis texensis («первоптица штата Техас»). Статус протоависа спорен. Некоторые палеонтологи (Е. Курочкин, Д. Петерз, Л. Мартин, Л. Хо) принимают птичий статус протоависа. Однако большинство палеонтологов отвергает связь протоависа с птицами и подвергает сомнению законность таксона. Согласно реконструкции Чаттерджи длина протоависа составляла около 35 см. Хотя он жил за 60-75 млн лет до археоптерикса, строение его скелета ближе к современным птицам. Из-за наличия зубов на клюве в нём предполагают хищника, размещение глаз на передней части черепа свидетельствует о ночном или сумеречном образе жизни. По мнению Чаттерджи, Протоавис был адаптирован к полету, на его костях присутствуют особые отметины — точки прикрепления перьев. Однако, по мнению других палеонтологов, небольшое количество окаменелостей не позволяет определить степень способности протоависа к полёту, более того, детальный их анализ не даёт никаких свидетельств наличия у него перьев. Е. Курочкин выделяет следующие особенности, связывающие протоависа с птицами.
Окаменелости протоависа найдены в 1984 в отложениях триасовой речной дельты. Они были смешаны с костями различных динозавров, что является возможным следствием массовой гибели при наводнении. Обнаруживший их учёный Шанкр Чаттерджи убеждён, что найденные им останки принадлежат двум особям одного вида - молодой и старой. К ним относятся лишь череп и фрагменты конечностей, достаточно диспропорциональные между собой, вследствие чего и появилась версия о химерности экземпляров, иными словами, гипотеза о том, что реконструированные скелеты, на самом деле, представляют собой скопление костей нескольких животных, принадлежащих к разным видам. Из-за небольшого числа находок и обстоятельств их обнаружения среди палеонтологов сохраняются сомнения относительно того, является ли протоавис птицей, и даже того, является ли он реально существовавшим животным. Российский палеорнитолог Е. Н. Курочкин, принимавший участие в изучении останков, полагает, что причина критического отношения учёных к протоавису носит преимущественно психологический характер, связанный с категоричностью утверждений Чаттерджи, который настаивает на способности обнаруженного существа к полёту. Принятие птичьего статуса протоависа подразумевает, что становление класса птиц произошло не позднее конца триасового периода. Следовательно, предков птиц следует искать среди триасовых архозавров. Это не согласуется с наиболее распространённой точкой зрения на филогению ранних птиц, согласно которой самыми близкими родственниками птиц являются троодонтиды и дромеозавриды — два семейства хищных динозавров (тероподов). Если данная филогенетическая теория верна, то эти семейства должны были возникнуть ещё в триасовом периоде, однако наиболее ранние окаменелости представителей этих двух семейств относятся к раннему мелу. По мнению Чаттерджи, ближайшими родственниками птиц являются триасовые цератозавры — группа тероподных динозавров, обладавшая рядом существенных особенностей, характерных для птиц (близкое к гетероцильному строение шейных позвонков, слияние тазовых костей и др.). Эти особенности скелета цератозавров традиционно расцениваются как результат конвергенции. Гипотеза дифилетического происхождения птиц, разрабатываемая Е. Н. Курочкином, частично основывается на изучении протоависа. Согласно этой гипотезе, веерохвостые птицы произошли от некоторой группы юрских архозавров, близким родственником которых является протоавис. Археоптерикс и энанциорнисы, согласно данной гипотезе, в свою очередь, являются потомками тероподов. Признание протоависа наиболее ранней на сегодняшний день птицей порождает временной промежуток в 85 млн лет между протоависом (210 млн лет) и первыми бесспорными веерохвостыми птицами (125 млн лет). В течение этих 85 млн лет (поздний триас, юрский период, ранний мел) должны были существовать примитивные птицы, окаменелостей которых однако не найдено. Исключение составляет археоптерикс (поздний юрский период), но он менее продвинут по сравнению с предполагаемой анатомией протоависа. Л. Уитмер заключил, что череп протоависа на самом деле принадлежит целурозавру. Эта оценка предполагает, что целурозавры появились не позднее конца триаса. Американские палеонтологи во главе с С. Чаттерджи (S. Chatterjee) при раскопках в районе Поста (штат Техас) обнаружили останки двух ископаемых позвоночных возрастом около 225 млн лет. Изучив находку, исследователи пришли к выводу, что имеют дело с наиболее древней среди известных науке птиц, жившей за 75 млн лет до археоптерикса, который, как известно, долгое время считался древнейшей первоптицей. Фрагментарность останков затрудняет определение и детальное описание находки, получившей название Protoavis. Ясно, однако, что эта предптица, как и археоптерикс, обладала когтистыми пальцами, хвостом и зубами, а следовательно, была близка к динозаврам. С другой стороны, в скелете протоависа найдена «вилочка» — грудная кость, встречающаяся только у настоящих птиц и отсутствовавшая у археоптерикса. В связи с этим Чаттерджи полагает, что протоавис обладал большей способностью к полету, чем археоптерикс; подкрепляется это предположение и строением костей крыла протоависа. Его череп весьма сходен с черепом современных птиц, а в задней части челюстей зубы отсутствуют. Возможно, протоавис стоял в начале эволюционной линии, приведшей древних птиц к постепенному избавлению от тяжелой, несущей зубы, челюсти и к способности свободного полета. Это предположение, однако, не полностью разделяет крупнейший палеонтолог и специалист по проблематике происхождения птиц Дж. Остром (J. Ostrom; Йельский университет, Нью-Хейвен, США), полагая, что имеющихся данных недостаточно, чтобы судить о столь значительной эволюционной «продвинутости» протоависа. С другой стороны, многие палеонтологи и раньше обращали внимание на то, что археоптерикс слишком примитивен и вместе с тем специализирован, чтобы служить прямым предком птиц. Кроме того, для этого ему «отпущено» слишком мало времени: ведь всего несколько миллионов лет спустя уже существовали настоящие птицы. Рядом с техасской находкой отпечатков оперения не обнаружено, однако на локтевых костях заметны выступы, к которым могли бы прикрепляться перья. Останки ископаемых птиц — вообще большая редкость, так как их хрупкие кости обычно плохо сохраняются. В данном случае обе найденные особи, вероятно, погибли при паводке или наводнении и их останки оказались захоронены под слоем ила. Chatterjee was convinced that some of these crushed bones belonged to two individuals – one old, one young – of the same species. However, only a few parts were found, primarily a skull and some limb bones which moreover do not well agree in their proportions respective to each other, and this has led many to believe that the Protoavis fossil is chimaeric, made up of more than one organism: the pieces of skull appear like those of a coelurosaur, while most parts of the limb skeleton suggest affinities to ceratosaurs and at least some vertebrae are most similar to those of Megalancosaurus, an avicephalan diapsid. If it really is a single animal and not a chimera, Protoavis would raise interesting questions about when birds began to diverge from the dinosaurs, but until better evidence is produced, the animal's status currently remains uncertain. Furthermore, palaeobiogeography suggests that true birds did not colonize the Americas until the Cretaceous; the most primitive lineages of unequivocal birds found to date are all Eurasian. Certainly, the fossils are most parsimoniously attributed to primitive dinosaurian and other reptiles as outlined above. However, coelurosaurs and ceratosaurs are in any case not too distantly related to the ancestors of birds and in some aspects of the skeleton not unlike them, explaining how their fossils could be mistaken as avian. Palaeontologist Zhonghe Zhou stated: "[Protoavis] has neither been widely accepted nor seriously considered as a Triassic bird ... [Witmer], who has examined the material and is one of the few workers to have seriously considered Chatterjee’s proposal, argued that the avian status of P. texensis is probably not as clear as generally portrayed by Chatterjee, and further recommended minimization of the role that Protoavis plays in the discussion of avian ancestry." Welman has argued that the quadrate of Protoavis displays synapomorphies of Theropoda. Paul has demonstrated the drepanosaur affinities of the cervical vertebrae. Gauthier & Rowe, and Dingus & Rowe have argued convincingly for identifying the hind limb of Protoavis as belonging to a ceratosaur. Feduccia has argued that Protoavis represents an arboreal "thecodont". In a study of early ornithischian dinosaurs, Sterling Nesbitt and others determined some of the partial remains of Protoavis to be a non-tetanuran theropod. The entire skull and neck are considered to be most likely from a drepanosaurid because the skull and neck are too big compared to the dorsal vertebrate of Protoavis.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| Time | Event |
| 09:18 pm [industrialterro] [Link] |
Adelobasileus Аделобазилевс (Adelobasileus, от др.-греч. ἄδηλος βασιλεύς «неясный царь») — вымершее протомлекопитающее позднего триасового периода (ок. 225 млн лет назад). Хищник, размером приблизительно с мышь. Известен по одному черепу, найденному на территории Техаса в 1989 году палеонтологами из Музея естественной истории и науки Нью-Мексико, где и хранится находка. Особенности строения черепа, особенно устройство внутреннего уха, свидетельствуют о том, что аделобазилевс был переходной формой между цинодонтами и триасовыми млекопитающими. Аделобазилевс жил приблизительно на 10 млн лет раньше наиболее продвинутых в эволюционном плане цинодонтов и всех других известных маммалиаформов (Mammaliaformes). Это даёт основания предполагать, что он мог быть общим предком млекопитающих, или, что более вероятно, состоял в близком родстве с общим предком млекопитающих.
Tags: Вымершие млекопитающие, Триас |
| Time | Event |
| 09:35 pm [industrialterro] [Link] |
Sinoconodon Синоконодон (Sinoconodon «китайский конический зуб») — вымершее животное, протомлекопитающее поздего триасового периода. Окаменелости найдены в Китае, провинция Юньнань, возраст — около 208 млн лет. Самым близким известным родственником синоконодона был, вероятно, морганукодон (возраст останков — 205 млн лет). Тем не менее, синоконодон существенно отличается от других маммалиаформов: его рост, по-видимому, не прекращался в течение всей жизни, точная окклюзия (смыкание зубов) отсутствует, что является примитивным признаком. Большинство зубов постоянно менялись, кроме передних щёчных, которые в случае утраты не восстанавливались и ни на одном известном черепе не сохранились. Синоконодон в целом считается более примитивным, обладает менее выраженными признаками млекопитающих, чем родственные ему маммалиаформы, включая морганоконудонтов и докодонтов, которые появились через несколько миллионов лет, и, вероятно, является реликтом более ранней стадии эволюции млекопитающих. В связи с этим, в одних работах он упоминается как самое примитивное млекопитающее, в других вовсе не считается млекопитающим. Синоконодон (Sinoconodon Patterson и Olson, 1961) – позднетриасовое млекопитающее примерно 15 см длиной. Вероятно, имел маленькое, пушистое, подобное полёвке тело с относительно крупной головой (приблизительно 60 мм в длину и 23 мм в ширину) с большими глазами и рептильными зубами, постоянно сменяющимися на протяжении всей жизни животного. Sinoconodon обладал примитивными "триконодонтными” щечными зубами, приспособленными к хищническому или насекомоядному образу жизни. К сожалению, постчерепные останки не позволяют определить локомотивные особенности передвижения. Sinoconodon имел широкий онтогенетический диапазон изменения размеров черепа: наименьший известный череп приблизительно 20 мм длиной, а наибольший - крупнее, чем 60 мм. Постчерепные окаменелости Sinoconodon демонстрируют, что у него имелись развитые передние конечности и плечевой пояс, мало чем отличающиеся от таковых у цинодонтов (Jenkins, 1971; Kemp, 1982; Sues, 1983), и современных монотремат. Sinoconodon однозначно помещен в Mammalia, потому что разделяет много особенностей с другими млекопитающими - например, краниомандибулярный сустав, сформированный дентарным мыщелком и гленоидом сквамозной кости, мысок височной кости, её антериорная стенка и костистая впадина для узла тройничного нерва. Однако расположение зубов Sinoconodon примитивно. Sinoconodontidae сохранили примитивное чередование и многократные замены резцов и клыков, замеченных у многих цинодонтов. Со временем, антериорные постклыковые зубы последовательно выпадали, приводя к постепенно увеличивающейся постклыковой диастеме, идентичной таковой у гомфодонтных цинодонтид (Hopson, 1971). Нижнечелюстные структуры и структуры среднего уха - по существу те же самые, как у Morganucodon и Megazostrodon, с примитивной постдентарной ложбинкой (хотя более уменьшенной, чем у Morganucodon).
Crompton и Sun (1985) наглядно показали, что верхние и нижние постклыковые зубы Sinoconodon отличаются по размеру и что они имеют переменное число. Верхние и нижние постклыковые зубы не совмещаются, так как на их выступах нет никаких зон изнашивания. Это - примитивная особенность, которая принудила Crompton и Sun (1985) предположить, что Sinoconodon более примитивен, чем многие из родственных ему млекопитающих: Morganucodon, Megazostrodon, Kuehneotherium, докодонты и харамивиды. Основываясь на дополнительных характерах зубной замены, краниомандибулярного сустава и ушной области, Crompton и Luo (1993) предложили, что Sinoconodon должен являться сестринским таксоном по отношению к другим группам млекопитающих. Таким образом, Sinoconodontidae представляют одно из ранних боковых происхождений позднетриасовых-раннеюрских млекопитающих.
Млекопитающие имеют два апоморфия слухового региона: удлиненный улитковый канал и возвышенность на тимпанной стороне кохлеарной области, именуемого как мысок верхнего края пирамиды височной кости. У цинодонтов улитка короче, меньше и сориентирована более медиально, чем улитковый канал млекопитающих; их кохлеарное размещение сформировано предушной областью и базисфеноидом. Ископаемые материалы Adelobasileus и Sinoconodon подтверждают развитие у ранних млекопитающих кохлеарной области. Sinoconodon имеет полностью сформированный мысок верхнего края височной кости, но короткую улитку, которая расширена только на одну треть длины мыска. Кохлеарный канал Morganucodon – вдвое длиннее, чем у Sinoconodon как относительно черепа, так и мыска. Более поздние млекопитающие - как, например, Haldanodon и мультитуберкуляты, демонстрируют более длинный улитковый канал. Предполагается, что мысок, преобразованный расширением височной кости за счет базисфеноида (вероятно, базиокципитально) является свидетельством перехода цинодонтов в млекопитающие (Luo, Z.-X., A.W. Crompton, и S.G. Lucas, 1995). Это изменение в окостенении и ориентации кохлеарной области эффективно для лучшей акустической изоляции внутреннего уха.
Зубная формула: 5.1.2.5/5.1.2.5. Все типы зубов являются вариабельными и нумеруются вследствие цинодонтоподобных непрерывных зубных замен в корреляции с ростом черепа. Sinoconodontidae сохранили переменные замены резцов и клыков, замеченные у большинства цинодонтов. Клыки заменялись, по крайней мере, четыре раза. Размер функциональных клыков является очень переменным среди различных черепных останков (Crompton и Luo, 1993). В самой полной форме клык подобен кинжалу. Верхний клык является настолько большим, что достигает нижней границы нижней челюсти, но на ранней стадии роста коронка клыка может быть короткой и конической.
Фактическое число функционирующих резцов является переменным. У некоторых черепных останков наличествовало только три функциональных резца в результате продолжающихся замен. Резцы имеют большие и небольшие размеры. Переменное расположение маленьких и больших зубов типично для цинодонтов, имевших непрерывную и переменную замену. Премоляры только присутствуют в ювенильных экземплярах (Zhang и др., 1998). Первый премоляр, кажется, потерян без замены у старших экземпляров. Второй премоляр имеет листообразную форму у молодых особей, и впоследствии исчезает у более старших возрастов. Нижние постклыковые зубы вообще "триконодонтоподобны", но демонстрируют более широкий диапазон морфологического изменения, возможно связанного с их заменой. У предпоследних моляров некоторых больших черепов имеется срединный цингулидный бугорок, который отсутствует у моляриморфного зуба меньших черепов. Этот выступ является переменной особенностью и может отсутствовать в антериорных постклыковых зубах. У верхнего моляра меньших экземпляров есть "триконодонтоподобная" трёхвершинная коронка с острыми выступами. В отличие от этого, верхний моляр больших экземпляров имеет тенденцию развивать дополнительный дистальный бугор. Как упомянуто ранее, у Sinoconodon вследствие вариаций в зубных размерах и морфологии между верхними и нижними постклыковыми зубами окклюзии нет.
Sinoconodon обладает многими характерами, предполагающими, что это - основная форма, относящаяся к Mammalia. Дальнейшие экземпляры (Crompton и Luo, 1993) в достаточной мере подтвердили это заключение. Подтверждение его статуса обеспечено дентальным мыщелком и гленоидной ямкой сквамозной кости, полнотой постгленоидного гребня и расширением черепа. В качестве мозаичных характеристик можно указать диапазон примитивных характеров, имеющихся у тритилодонтид и трителодонтов. Самым важным является расположение зубов - сравнение ряда экземпляров отличающихся размеров показывает, что способ замены зубов был аналогичен цинодонтам, а не млекопитающим (Zhang и др. 1998; Kielan-Jaworowska и др. 2004). Резцы и клыки сменялись несколько раз, в то время как антериорные премоляры выпадали, но не заменялись.
Филогенетично Sinoconodon был, несомненно, близок к основной, относящейся к млекопитающим радиативной линии, сохранив примитивные характеры. Действительно, не принимая во внимание малоизвестного Adelobasileus и харамивид, он универсально принят как сестринская группа остальной части Mammalia. Как и у морганукодонтов, между орбитами Sinoconodon есть костистое разделение и сустав между сквамозной и зубной костями, но они более уменьшены, чем у морганукодонтид (Crompton и Sun, 1985). Однако, Sinoconodon, кажется, более примитивен, чем Morganucodon. Кохлеарная структура была более примитивной (Luo и др., 1995), и отсутствие последовательных зон изнашивания указывает, что Sinoconodon испытывал недостаток в смыкании коренных зубов. Это напоминает цинодонтов в сохранении большой септомаксиллярной кости и многократной замене резцов и клыков. Данные особенности предполагают, что Sinoconodon отклонился от относящегося к млекопитающим эволюционного ствола ранее, чем морганукодонтиды.
 Tags: Вымершие млекопитающие, Триас |























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Previous Day | 2012/03/27 [Archive] |
Next Day |