[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 20 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| October 23rd, 2014 | |
|---|---|
| 12:42 pm [industrialterro] [Link] |
Pterodaustro Pterodaustro is a genus of Cretaceous pterodactyloid pterosaur from South America, which lived 105 million years ago. The genus was named in 1969 by José Bonaparte as an as yet undescribed nomen nudum. The first description followed in 1970, making the name valid, the type species being Pterodaustro guiñazui. The genus name is derived from Greek pteron, "wing" and Latin auster, "south (wind)". The elements are combined as a condensed pteron de austro, "wing from the south". The specific name honours paleontologist Román Guiñazú. It was emended in 1978 by Peter Wellnhofer into guinazui, because diacritical signs such as the tilde are not allowed in species names. The back of the skull was also rather elongated and in a low position; there are some indications for a low parietal crest. Pterodaustro had an adult wingspan of approximately 250 centimetres (8.20 ft). Its hindlimbs are rather robust and its feet large. Its tail is uniquely elongated for a pterodactyloid, containing 22 caudal vertebrae, whereas other members of this group have at most sixteen. Robert Bakker incorrectly suggested that, like flamingos, this pterosaur's diet may have resulted in a pink hue. However, recent studies show that only Neoaves can sequester carotenoids for use as a pigment in the feathers rather than just the skin or beak, and even then they require to enhance it with structural colors, so a pink hue for any pterosaur seems extremely unlikely. At least two specimens of Pterodaustro have been found, MIC V263 and MIC V243, with gizzard stones in the stomach cavity, the first ever reported for any pterosaur. These clusters of small stones with angled edges support the idea that Pterodaustro ate mainly small, hard-shelled aquatic crustaceans using filter-feeding. Such invertebrates are abundant in the sediment of the fossil site. A study of the growth stages of Pterodaustro concluded that juveniles grew relatively fast in their first two years, attaining about half of the adult size. Then they reached sexual maturity, growing at a slower rate for four to five years until there was a determinate growth stop. In 2004 a Pterodaustro embryo in an egg was reported, specimen MHIN-UNSL-GEO-V246. The egg was elongated, six centimetres long and 22 millimetres across and its mainly flexible shell was covered with a thin layer, 0.3 mm thick, of calcite. Three-dimensionally preserved eggs were reported in 2014. Comparisons between the scleral rings of Pterodaustro and modern birds and reptiles suggest that it may have been nocturnal, and may have had similar activity patterns to modern anseriform birds that feed at night. The genus Puntanipterus might be a subjective junior synonym of Pterodaustro. Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, ктенохазматоидеи, монофенестраты, птеродактили, птерозавры |
| October 20th, 2014 | |
| 09:02 pm [industrialterro] [Link] |
Pteranodon Птеранодон (от др.-греч. πτερόν — «крыло» и ἀνόδους — «беззубый») — род птерозавров. Известен с верхнего мела Северной Америки. Отличался крупным выростом на голове (выполнявшим роль руля при полёте) и беззубым клювом. Птеранодон был одним из крупнейших летающих ящеров, с размахом крыльев более 6 метров, обитавшим в меловом периоде в небесных просторах над территорией современной Северной Америки (находки в штатах Канзас, Алабама, Небраска, Вайоминг и Южная Дакота). Количество найденных окаменелостей этого птерозавра превышает 1 500 экземпляров – среди летающих рептилий это абсолютный рекорд. Причем окаменелости очень высокой степени сохранности и детализации. Бесспорно – эти существа имели огромные популяции на побережьях гигантского Западного внутреннего морского пути (Niobraran Sea – теплое море мелового периода, располагавшееся в центральной части Североамериканского континента от Мексиканского залива до Канады). Птеранодон в отличие от более ранних Rhamphorhynchus в клюве не имел зубов и тем самым походил на современных птиц. Однако края челюстей у него были тонкие и острые, образованные особо твердой костной тканью. Верхняя челюсть длиннее нижней и изогнута вверх, причем некоторые подвиды имеют достаточно большой уровень восходящего изгиба, одновременно с расширением клюва к концу (Drawndraco, 2010). Другими характеристиками, которые отличают птеранодонов от других птерозавров, являются узкие отростки спинных позвонков, пластинчатые костистые связки, которыми крепились позвонки выше бедер, и относительно короткий хвост, в котором последние несколько позвонков сливаются в длинный стержень. Хвост составлял около 3,5% длины птеранодона, т.е. около 25 см от длины взрослого самца. Наиболее отличительная черта – черепной гребень, т.е. удлиненные плоские лобные кости ящера, выступающие немного вверх и назад. Причем размер и форма этих гребней менялась в зависимости от возраста и пола птерозавра. Pteranodon sternbergi (Geosternbergia) – один из наиболее ранних видов, самцы которого имели наиболее крупные и широкие гребни, которые загнуты более вертикально, нежели у их потомков – Pteranodon longiceps, которые были обладателями более узкого и вытянутого гребня. Функции этого гребя – вопрос достаточно дискуссионный, однако тот факт, что у самок он был значительно меньше, говорит о социальном значении этого выроста; однако некоторые специалисты (Беннет С., 1992 г.) говорят о «летательных» характеристиках гребня, который уравновешивал (стабилизировал) при полете длинный клюв ящера. Еще в 1910 г. Джордж Фрэнсис Итон высказывался о строго аэродинамических характеристиках и назначении гребня птеранодона, который выступал по его мнению в качестве противовеса и массивной точки крепления мощных летательных мышц и мышц челюстей. Однако тот факт, что размер гребня увеличивался с возрастом ящера и имел широкие вариации в размерах в рамках даже одной популяции говорит о том, что его аэродинамические свойства все-таки вторичны. Беннет С. в 1992 г. согласился с указанными доводами и пояснил, что основной вторичной функцией гребня выступал именно противовес как освобождение шеи от чрезмерной нагрузки на мышцы, необходимой для ориентации клюва. Тем не менее, в расчет принимались только размерные классы гребней, принадлежащие мужским особям птеранодона. У самок гребни были намного меньше, слегка округлой формы. Причем испытания, проведенные в аэродинамической трубе, показали, что мышечные нагрузки, приходящиеся на гребень, при поворотах головы из стороны в сторону – были гораздо больше, нежели те, которые были необходимы для стабилизации клюва при полете. То есть ящер был способен уравновесить свой клюв и без помощи гребня. С учетом вышеуказанных доводов, мнения специалистов переключились на обоснование функции гребня как рулевого органа. Еще в 1943 г. Доминик фон Крипп и Росс С. Штейн говорили о том, что между гребнем и спиной у этого ящера возможно даже был натянут кожистый парус, и опять же эта теория не принимала в расчет самок. Кроме того, Беннет С. обратил внимание на то, что гребень-руль не смог бы обеспечить хоть сколько-нибудь эффективную маневренность в полете, которая очень просто достигается путем изменения натяжения крыла. Поэтому основная функция гребня птеранодона, по все видимости, ограничивается социальным и половым поведением. Эти летающие ящеры имели ярко выраженный половой диморфизм – самки были меньше в размерах. Гребень у самок меньше и округлой формы, тазовые кости у самок значительно расширены, что является следствием откладывания яиц. Вполне вероятно, что, как и другие полигамные животные (у которых мужские особи соревнуются за спаривание с гаремами женских), птеранодоны жили в основном на морских лежбищах, где они могли бы гнездиться вдали от наземных хищников и питаться далеко от берега; больше всего их окаменелостей найдено в местах, которые в то время были в сотнях километров от береговой линии. Форма крыла птеранодона говорит о том, что передвигался в воздухе он наподобие современных альбатросов, поскольку имеет такое же соотношение длины крыла и длины позвоночника: 8 к 1. Этот способ полета получил название "динамическое парение", т.к. альбатрос (как и птеранодон) может использовать восходящие потоки, формирующиеся у поверхности океана и преодолевать десятки километров без единого взмаха крыла. Все птеранодоны делятся на две группы. Первая была меньше по размеру, имела небольшие закругленные гребни и широкие тазовые кости; размах крыльев этой группы был не более 4-х метров. Широкие тазовые кости вполне однозначно указывают на самок. Самцы были гораздо крупнее и могли иметь размах крыльев, вероятно, свыше 7 метров. Вес птеранодонов оценивался разными учеными по-разному и привел к крайне расходящимся результатам в диапазоне от 20 до 93 кг. Тем не менее, в 2010 г. были проведены детальные исследования (М. Уиттон), которые показали, что по своим «прочностным» характеристикам крыло птеранодона было способно и на эффективный «машущий» полет над поверхностью земли, вопреки укоренившемуся мнению, что эти ящеры только парили в прибрежных водах. Это же исследование показало, что птеранодон вполне был способен подниматься в воздух и с четвероногой позиции – он опирался на мощные передние конечности (как отжимание в упоре лежа), совершал небольшой прыжок и потом – быстрое вторичное движение машущими крыльями, которое уже уносило ящера в воздух. Исследования дорожек следов, которые были открыты в 1974, 1994 и 2001 годах (Беннет С., Брамвелл и Уитфилд) позволяют сказать, что птеранодон перемещался во земле в четвероногой позиции, разместив свернутую перепонку крыла позади себя. Пищевой рацион ящера составляла в основном рыба, об этом свидетельствуют находки окаменелостей рыб в желудках и между челюстей птеранодонов. Традиционно способ охоты птеранодона представляют как низкое парение над водой и периодическое погружение клюва вслед замеченной добыче. Это предположение (Беннет С. 1994 г.) было основано на том, что ящер не мог взлетать с поверхности воды. Однако исследования М. Уиттона показали, что птеранодон мог использовать механизм взлета с воды, такой же, как и с поверхности земли. Поэтому охотничья тактика этого ящера могла быть очень вариативной: вплоть до пикирующего погружения в океан, как это делают современные североатлантические олуши, причем «заточенный» профиль Птеранодона, массивный гребень и возможность складывать крылья за спиной – это весьма эффективные эволюционные преимущества, которые позволяли этому ящеру быть весьма успешным при таком виде охоты. ( Read More ) Ископаемые останки и реплики (1, 2, 3, 4, 5, 6, 7, 8, 9): ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеранодонтиды, птеранодонтойды, птеродактили, птерозавры |
| October 11th, 2014 | |
| 02:40 pm [industrialterro] [Link] |
Phosphatodraco Phosphatodraco (meaning "phosphate dragon", in reference to the phosphates of Morocco, the country where it was found) is a genus of azhdarchid pterodactyloid pterosaur from a late Maastrichtian-age Upper Cretaceous portion of the Oulad (or Qualad) Abdoun Phosphatic Basin, Grand Doui, near Khouribga, central Morocco. Phosphatodraco is based on holotype OCP DEK/GE 111, found in 2000, which is composed of five associated, though disarticulated and compressed, damaged cervical vertebrae and a bone of unknown origin. The cervical vertebrae are thought to be a series from the fifth (the longest with a length of thirty centimeters) to the ninth. The individual to which the neck belonged would have had a wingspan of about five meters (16.4 feet). It is unusual among azhdarchids for having elongate vertebrae at the base of the neck (also with neural spines), interpreted as modified dorsal vertebrae; the neck is also one of the most complete known for azhdarchids. It was one of the last pterosaurs before the Cretaceous–Paleogene extinction event that finished off the group, and is the first azhdarchid found in northern Africa. P. mauritanicus is the type and only known species. The specific name refers to Mauretania.    Вместе с саркозухом. Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархиды, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| 02:01 pm [industrialterro] [Link] |
Plataleorhynchus Plataleorhynchus was a genus of ctenochasmatid pterodactyloid pterosaur from the Tithonian-Berriasian-age Upper Jurassic or Lower Cretaceous Purbeck Limestone of Dorset, England. The genus was named in 1995 by Stafford Howse and Andrew Milner. The type species is Plataleorhynchus streptophorodon. The genus name is derived from Platalea, the spoonbill, and Greek rhynchos, "snout", in reference to the distinctive form of the front of the skull. The specific name is derived from Greek streptophoros, "collared", and odon, "tooth", referring to the tooth form. It is based on holotype NHML R.11957 (earlier BMNH R.11957), an incomplete anterior upper jaw with teeth found in a chalkstone quarry near Langton Matravers. The fossil is present on a plate; its underside is visible. This jaw is notable because it expands to form a circular, spatula-like shape at the front, holding 22 narrow teeth that point sideways. Forty other teeth (sockets) were present in the preserved remainder of the snout; the total for the upper jaws was estimated at 76. The authors classified Plataleorhynchus as a member of the Ctenochasmatidae, a group containing many filter feeders. David Unwin in 2005 placed it in the subgroup of the Gnathosaurinae. Although Plataleorhychus would have been similar in size to large gnathosaurines like Gnathosaurus; its skull length was estimated at a minimum of forty centimetres (15.75 in), the different shape of its spoonbill, presence of an apparently horn-covered pad on the palate, and smaller teeth suggest it did not feed in the same way, perhaps stirring up water-dwelling animals from muddy or weedy environments. Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, ктенохазматоидеи, монофенестраты, птеродактили, птерозавры |
| October 9th, 2014 | |
| 12:50 pm [industrialterro] [Link] |
Ornithocheirus Орнитохейр (др.-греч. ὄρνις «птица» и χείρ «рука») — крупный птерозавр раннего мелового периода с размахом крыльев в 2,5 - 5 метров. Орнитохейр обитал в Европе 110 млн. лет назад. Его узкий удлинённый череп плавно переходил в длинный клюв, унизанный острыми зубами. Орнитохейры, по-видимому, населяли побережья морей, берега больших озёр и речные поймы.
Используя восходящие потоки воздуха, наиболее крупные орнитохейры вполне могли пролетать сотни километров, едва взмахивая крыльями. Этому способствовал также относительно небольшой вес, поскольку в костях рептилии находились так называемые «воздушные мешки». С высоты эти птерозавры опускались к поверхности воды за рыбой. Заметив добычу, птерозавр мгновенно погружал в воду клюв, смыкал челюсти и заглатывал рыбу целиком. Высокий килевидный гребень на клюве, по-видимому, помогал орнитохейру удерживать его в вертикальном положении в воде. На суше орнитохейр, как и большинство других птерозавров, двигался медленно и неуклюже, что делало его лёгкой добычей даже для некрупных хищников. Чтобы обезопасить себя от врагов, птерозавры гнездились многочисленными колониями недалеко от воды. А это значит, что прибрежные скалы, утёсы и другие наиболее безопасные и удобные места для гнездования были перенаселены. Окаменевшие кости орнитохейра впервые были найдены в Южной Англии в 1827 году, но свое название этот птерозавр получил только в 1869 году. Размах крыльев у большинства видов орнитохейра не превышал 2,5 м, однако сравнительно недавно в бразильских горах Сантана были найдены останки настоящего орнитохейра-исполина, размах крыльев которого превышал 8 м (ныне этот вид реклассифицирован в отдельный род Tropeognathus). Хотя окаменелости птерозавров известны ученым более 250 лет, одним из первых официального научного названия удостоился орнитохейр. Было описано около 30 видов орнитохейров, но ныне многие из них переклассифицированы в другие роды птерозавров. Ornithocheirus (from Greek "ορνις", meaning bird, and "χειρ", meaning hand) is a pterosaur genus known from fragmentary fossil remains uncovered from sediments in the UK. Several species have been referred to the genus, most of which are now considered as dubious species, or members of different genera, and the genus is now often considered to include only the type species, Ornithocheirus simus. Species have been referred to Ornithocheirus from the mid-Cretaceous period of both Europe and South America, but O. simus is known only from the UK. Because O. simus was originally named based on poorly preserved fossil material, the genus Ornithocheirus has suffered enduring problems of zoological nomenclature. Fossil remains of Ornithocheirus have been recovered mainly from the Cambridge Greensand of England, dating to the beginning of the Albian stage of the late Cretaceous period, about 110 million years ago. Additional fossils from theSantana Formation of Brazil, dating to 112-108 million years ago, are sometimes classified as species of Ornithocheirus, but have also been placed in their own genera, most notably Tropeognathus. The original material of Ornithocheirus simus, recovered from England, indicates a mid-sized species with a wing span of 2.5 m (8.2 ft). Referred specimens attributed to Ornithocheirus simus can reach 5 m (16.5 ft). O. simus bore a distinctive convex "keeled" crest on its snout. Unlike the related Anhanguera and Coloborhynchus, which had an expanded rosette of teeth at the jaw tips, Ornithocheirus species (including O. simus) had straight jaws that narrowed toward the tip. Also unlike related pterosaurs, the teeth of Ornithocheiruswere mostly vertical, rather than set at an outward-pointing angle. They also had fewer teeth than related species. The type specimen of Ornithocheirus simus is represented only by a broken piece of the upper jaw tip. While it does preserve several characteristic features of Ornithocheirus, it is nearly identical to comparable bones in Tropeognathus mesembrinus, making clear distinction between these two species impossible. ( Read More )</span> Репродукции (1, 2, 3, 4, 5, 6): Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейриды, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| 12:01 pm [industrialterro] [Link] |
Muzquizopteryx Muzquizopteryx is a genus of pterodactyloid pterosaur (flying reptile) from the Late Cretaceous of what is now Coahuila,Mexico. In the nineties José Martínez Vásquez, a worker at the chalk quarry of El Rosario, uncovered a skeleton of a pterosaur. This he handed to a quarry official, who had it bricked in on the face of an office wall as a decorative piece. After its unique scientific value had been recognised in 2002, the specimen was acquired by the Universidad Nacional Autónoma de México. Subsequently it was studied by a combined team of the University of Karlsruhe and the University of Heidelberg, in this financially supported by the Volkswagen company. The specimen was scientifically reported in 2004. In 2006 the type species Muzquizopteryx coahuilensis was named and described by Eberhard Frey, Marie-Céline Buchy,Wolfgang Stinnesbeck, Arturo González-González and Alfredo di Stefano. The generic name is derived from the Múzquizdistrict and a Greek πτέρυξ, pteryx, "wing". The specific name is derived from the state of Coahuila. Muzquizopteryx is based on holotype UNAM IGM 8621, found in the El Rosario layers, early Coniacian-aged rocks. It consists of a nearly complete, articulated skeleton that includes soft tissue remains, among them long fossilised tendons along both sides of both lower arms. The specimen represents an adult individual. In 2012 a second specimen was reported, MUDE CPC-494, again uncovered by a quarry worker, perhaps at the same site, and sold to a private collector. It was later acquired by the Museo del Desierto Saltillo. It consists of the right upper wing of a subadult individual, with about 81% of the length of the holotype. As its provenance probably consists of slightly older layers from the late Turonian and the remains are limited, it was referred to a Muzquizopteryx sp. Muzquizopteryx was relatively small for a pterodactyloid pterosaur, with a wingspan of around two metres (6.6 ft). It had an elongated head with a convex upper profile, ending at the back of the head in a backward pointing short rounded crest. The jaws were toothless. The arms were very robust with the humerus featuring a large hatchet-shaped deltopectoral crest, indicating a strong wing musculature. The pteroid bone was long and pointed towards the neck, supporting a flight membrane. Muzquizopteryx was by its describers assigned to the Nyctosauridae. It would then be the oldest known member of the group and the smallest known; indeed the smallest adult Late Cretaceous pterosaur discovered until 2006. As Nyctosaurus is sometimes included with the Pteranodontidae, Muzquizopteryx too might be considered a member of that group under some classifications. Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, никтозавриды, орнитохейройды, птеродактили, птерозавры |
| October 8th, 2014 | |
| 02:31 pm [industrialterro] [Link] |
Nyctosaurus Nyctosaurus is a genus of pterodactyloid pterosaur, the remains of which have been found in the Niobrara Formation of the mid-western United States, which, during the late Cretaceous Period, was covered in an extensive shallow sea. The genus Nyctosaurus has had numerous species referred to it, though how many of these may actually be valid requires further study. At least one species possessed an extraordinarily large antler-like cranial crest. Nyctosaurus was similar in anatomy to its close relative and contemporary, Pteranodon. It had relatively long wings, similar in shape to modern seabirds. However, it was smaller overall than Pteranodon, with an adult wingspan of 2 meters (6.6 ft) and a maximum weight of about 1.86 kg. The overall body length was 37 cm. Some specimens preserve a distinctive crest, at least 55 cm tall in old adults, relatively gigantic compared to the rest of the body and over three times the length of the head. The crest is composed of two long, grooved spars, one pointed upward and the other backward, arising from a common base projecting up and back from the back of the skull. The two spars were nearly equal in length, and both were nearly as long or longer than the total length of the body. The upward-pointing crest spar was at least 42 cm long (1.3 ft) and the backward-pointing spar was at least 32 cm long (1 ft). The jaws of Nyctosaurus were long and extremely pointed. The jaw tips were thin and needle sharp, and are often broken off in fossil specimens, giving the appearance that one jaw is longer than the other, though in life they were probably equal in length. Nyctosaurus is the only pterosaur to have lost its clawed "fingers", with the exception of the wing finger (of which however the fourth phalanx was lost), which is likely to have impaired its movement on the ground, leading scientists to conjecture that it spent almost all of its time on the wing and rarely landed. In particular, the lack of claws with which to grip surfaces would have made climbing or clinging to cliffs and tree trunks impossible for Nyctosaurus. Nyctosaurus, like its relative Pteranodon, appears to have grown very rapidly after hatching. Fully adult specimens are no larger than some immature specimens such as P 25026, indicating that Nyctosaurus went from hatching to adult size (with wingspans of 2 m or more) in under a year. Some sub-adult specimens have been preserved with their skulls in nearly pristine condition, and lack any trace of a head crest, indicating that the distinctively large crest only began to develop after the first year of life. The crest may have continued to grow more elaborate as the animal aged, though no studies have examined the age of the fully adult, large-crested specimens. These individuals may have been 5 or even 10 years old at the time of their deaths. Only five relatively complete Nyctosaurus skulls have been found. Of those, one is juvenile and does not possess a crest (specimen FMNH P 25026), and two are more mature and may show signs of having had a crest but are too badly crushed to say for sure (FHSM 2148 and CM 11422). Two specimens (KJ1 and KJ2) described in 2003, however, preserved an enormous double-pronged crest. A few scientists had initially hypothesized that this crest, which resembles an enormous antler, may have supported a skin "headsail" used for stability in flight. While there is no fossil evidence for such a sail, studies have shown that a membranous attachment to the bony crest would have imparted aerodynamic advantages. However, in the actual description of the fossils, paleontologist Christopher Bennett argued against the possibility of a membrane or soft tissue extension to the crest. Bennett noted that the edges of each prong were smooth and rounded, and showed no evidence for any soft-tissue attachment points. He also compared Nyctosaurus with large-crested tapejarids, which do preserve soft tissue extensions supported by prongs, and showed that in those species, the attachment points were obvious, with jagged edges where the transition from bone to soft tissue occurred. Bennett concluded that the crest was most likely used solely for display, citing similar structures in modern animals. The 2009 study by Xing and colleagues testing the aerodynamics of the giant crest with a "headsail" also tested the aerodynamics of the same crest with no sail, and found that it added no significant negative factors, so a crest with no headsail would not have hindered normal flight. It is more likely that the crest acted mainly for display, and that any aerodynamic effects it may have had were secondary. Bennett also argued that the crest was probably not a sexually dimorphic character, as in most crested pterosaurs, including the related Pteranodon, both sexes are crested and it is only the size and shape of the crest that differs. The apparently non-crested Nyctosaurus specimens therefore probably came from sub-adults. Sankar Chatterjee and R.J. Templin used estimates based on complete Nyctosaurus specimens to determine weight and total wing area, and to calculate its total wing loading. They also estimated its total available flight power based on estimated musculature. Using these calculations, they estimated the cruising speed of Nyctosaurus gracilis as 9.6 meters/second (34.5 kilometers/hour or 21.4 miles/hour). All known Nyctosaurus fossils come from the Smokey Hills Chalk of Kansas, part of the Niobrara Formation. Specifically, they are found only within a narrow zone characterised by the abundance of ammonite fossils belonging to the species Spinaptychus sternbergi. These limestone deposits were laid down during a marine regression of the Western Interior Seaway that lasted between 85 and 84.5 million years ago. Therefore, Nyctosaurus was a relatively short-lived species, unlike its relative Pteranodon, which is found throughout almost all of the Niobrara layers into the overlying Pierre Shale Formation, and existed between 88 and 80.5 million years ago. The ecosystem preserved in this zone was unique in its abundance of vertebrate life. Nyctosaurus shared the sky with the bird Ichthyornis and with Pteranodon longiceps, though the second Niobrara Pteranodon species, P. sternbergi, had disappeared from the fossil record by this point. In the waters of the Western Interior Seaway below swam mosasaurs (Clidastes, Ectenosaurus and Tylosaurus), the flightless diving bird Parahesperornis, and a wide variety of fish, including swordfish-like Protosphyraena, the predatory Xiphactinus and the shark Cretolamna. The first Nyctosaurus fossils were described in 1876 by Othniel Charles Marsh, based on fragmentary material, holotype YPM 1178, from the Smoky Hill River site in Kansas. Marsh referred the specimen to a species of his new genus Pteranodon, as Pteranodon gracilis. Later that year, Marsh reclassified the species in its own genus, which he named Nyctosaurus, meaning "night lizard" or "bat lizard", in reference to the wing structure somewhat paralleling those of bats. In 1881, Marsh incorrectly assumed the name was preoccupied and changed it into Nyctodactylus, which thus is now a junior synonym. In 1902,Samuel Wendell Williston described the most complete skeleton then known (P 25026) discovered in 1901 by H. T. Martin. In 1903, Williston named a second species, N. leptodactylus, but this is today considered identical to N. gracilis. In 1953, Llewellyn Ivor Price named a partial humerus, DGM 238-R found in Brazil, N. lamegoi; the specific name honours the geologist Alberto Ribeiro Lamego. This species has an estimated wingspan of four metres; it is today generally considered to be a form different from Nyctosaurus, but has not yet been assigned its own genus name. In 1972 a new skeleton, FHSM VP-2148, in 1962 discovered by George Fryer Sternberg, was named N. bonneri; it is today generally seen as identical to N. gracilis. In 1978 Gregory Brown prepared the most complete Nyctosaurus skeleton currently known, UNSM 93000. In 1984 Robert Milton Schoch renamed Pteranodon nanus (Marsh 1881), "the dwarf", Nyctosaurus nanus. The question of this species validity is currently pending further study. In the early 2000s, Kenneth Jenkins of Ellis, Kansas collected two specimens of Nyctosaurus which were the first to demonstrate conclusively that not only was this species crested, the crest in mature specimens was very large and elaborate. The specimens were purchased by a private collector in Austin, Texas. Despite being in private hands rather than a museum collection, paleontologist Chris Bennett was able to study the specimens and gave them the manuscript reference numbers KJ1 and KJ2 (for Kenneth Jenkins). Bennett published a description of the specimens in 2003. Despite the unusual crests, the specimens were otherwise indistinguishable from other specimens of Nyctosaurus. However, the then-currently named species were extremely similar and Bennett declined to refer them to a specific one pending further study of the differences, or lack therof, between species of Nyctosaurus. ( Read More )       ( Read More )       Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, никтозавриды, орнитохейройды, птеродактили, птерозавры |
| October 1st, 2014 | |
| 03:05 pm [industrialterro] [Link] |
Nurhachius Nurhachius was a genus of istiodactylid pterodactyloid pterosaur from the Barremian-Aptian-age Lower Cretaceous Jiufotang Formation of Chaoyang, Liaoning, China. The genus was named in 2005 by Wang Xiaolin, Alexander Kellner, Zhou Zhonge and Diogenes de Almeida Campos. The type species is Nurhachius ignaciobritoi. The genus name refers to Nurhaci, the first khan of the Qing Dynasty, whose original power base encompassed the region where the fossil was found. The specific name honours the late Brazilian paleontologist Ignácio Aureliano Machado Brito, who pioneered the study of pterosaurs in his country. The genus is based on holotype IVPP V-13288, a partial skull and skeleton. The skull is elongated with a preserved length of 315 millimetres and an estimated total length of about 330 millimetres. The nasoantorbital fenestra, the large skull opening, is relatively long at 58% of the skull length. The lower jaws have a length of 291 millimetres. The skull is similar to that of Istiodactylus, which lived at about the same time in what is now England, especially in the teeth that are compressed side to side and the long fenestra. However, it differs from Istiodactylus in several details, including a significantly lower skull, different jugal and a slight curve to the upper margin of the lower jaw. The teeth are curved to the back, have three roots and are robust. They are limited to the anterior ends of the jaws; there are 28 teeth in the upper jaws and 26 in the lower jaws for a total of 54. Most elements of the postcranial skeleton are known, with the exception of some cervical vertebrae, the ribs, the tail and the two most extreme phalanges of the wing finger. Its wingspan was estimated at 2.4-2.5 m (7.87-8.20 ft). The describers pointed out several similarities with the Pteranodontoidea (here used in the sense of the clade Pteranodon + Istiodactylus + Anhangueridae) and one, the warped deltopectoral crest of the humerus, unique to just the clade Istiodactylus + Anhangueridae. A cladistic analysis by Wang showed Nurhachius to be a member of the Istiodactylidae within the much larger Dsungaripteroidea sensu Kellner. Its position as being closely related to Istiodactylus was supported by the analysis in a publication by Lü Junchang. In 2008 Lü suggested Nurhachius was a subjective junior synonym of Liaoxipterus. В Ляонине, северо-восточной области Китая, палеонтологи обнаружили окаменелости двух новых видов птерозавров.Feilongus youngi и Nurhachius ignaciobritoi разделяли небеса с ранними птицами 120 миллионов лет назад. Feilongus имел два гребня наверху головы, бегущих от кончика "носа" до её задней части. Один гребень — в передней части морды, другой — в задней части головы. Этот птерозавр имел неправильный прикус, а его зубы были изогнутыми и иглообразными. А вот зубы Nurhachius ignaciobritoi были треугольными.Оба вида принадлежат к группе, ранее найденной только в Европе. Размах крыльев, затянутых тонкой кожей, у этих летающих ящеров составлял около 2,5 метров. Учёные предполагают, что этим видам был свойственен не машущий полёт, а парение. Один из палеонтологов, опубликовавших новое большое исследование птерозавров, Александр Келлнер (Alexander Kellner) из федерального университета в Рио (Universidade Federal do Rio de Janeiro), сообщил любопытную подробность. Он отметил, что пересекавшиеся во времени птерозавры и ранние птицы населяли, в массе своей, различные среды обитания и очень мало конкурировали друг с другом. Птерозавры преобладали над птицами в прибрежных районах, в то время как птицы господствовали в глубине континента. Хотя и те, и другие, в принципе, встречались в каждом из этих мест.    Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, истиодактилиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| September 29th, 2014 | |
| 10:03 pm [industrialterro] [Link] |
Noripterus The first, holotype specimen of Noripterus (IVPP V.4062, type locality IVPP 64045) preserved the front part of the skull and lower jaws, vertebrae, and partial limbs and pelvis. Noripterus was quite similar to the contemporaneous Dsungaripterus, though it was estimated to be a third shorter. It has long narrow neck vertebrae and, like Dsungaripterus, a crest and no teeth in the front of the lower jaw. The teeth that are present are well-developed and spaced fairly far apart. The sharp snout is straight and not pointed upwards as with Dsungaripterus. Because of its similarity to Dsungaripterus, Noripterus has been assigned to the family Dsungaripteridae. The genus Phobetor, named after the Greek god of nightmares, was in 1982 originally described by Natasha Bakhurina as a species of Dsungaripterus (D. parvus), based on a single lower leg bone, PIN 3953. The discovery of more remains later, among which an almost complete skull, GIN 100/31, was reason for Bakhurina to name D. parvus in 1986 as a separate genus, and the species name became Phobetor parvus. However, the genus name Phobetor was already being used as a junior synonym of a species of sculpin, namely, the arctic staghorn sculpin, Gymnocanthus tricuspis (synonym "Phobetor tricuspis" Krøyer, 1844) and thus unavailable. In 2009, Lü and colleagues re-examined much of the known dsungaripterid fossil material, and found that "Phobetor" was indistinguishable from Noripterus, causing them to refer to it as a junior synonym. Assigning the "Phobetor" material to Noripterus increases the known size of the latter as it indicates a maximum wingspan of 4 metres (13.1 ft). Dsungaripterids like Noripterus are interpreted as adapted for feeding on fish and shellfish, with long narrow toothless beak tips for probing for and picking up suitable prey, and robust teeth farther back for cracking shells. The skulls of these animals are more robust than those of other pterosaurs, as well as their limbs and vertebrae. Noripterus lived in the same time and place as the larger Dsungaripterus, in formations that indicate the presence of extensive inland lake systems. Because Noripterus had a more lightly built skull with weaker, more slender teeth than its larger contemporary, it is likely that the two pterosaurs occupied separate ecological niches, with Dsungaripterus hunting in the shallow parts of lakes and eating hard-shelled animals, while Noripterus fed on fish from deeper regions of the lakes. Новый ископаемый материал птерозавра нориптеруса (Noripterus) из семейства джунгариптерид найден в местечке Tatal в западной Монголии (возраст ранний мел). Его останки позволили по новому диагностировать семейство Dsungaripteridae и нориптеруса. Все птерозавры, найденные в этом месте принадлежат джунгариптеридам (или джунгароптериксу или нориптериксу). Фобетор (Phobetor) представляет младший синоним нориптеруса. Различные формы передних половин черепов, а также зубов, говорят о том, что джунгариптерус и нориптерус могли занимать разные экологические ниши. Джунгариптерус имел размах крыльев около 5 метров, в то время как нориптерус - не больше 4 метров. У первого зубы толстые и мощные, позволявшие питаться не только рыбой, но и относительно крупными животными, а у второго зубы мелкие и слабые, тонкие и предназначенные только для рыбы. Джунгариптерус питался по берегам водоёмов, а нориптерус охотился преимущественно над глубокими водами.  Внизу, окрашен фиолетовым цветом.    Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, джунгариптероиды, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| 09:32 pm [industrialterro] [Link] |
Ningchengopterus Ningchengopterus is a genus of pterodactyloid pterosaur from the Early Cretaceous-age Yixian Formation of China. It is known from a juvenile specimen, holotype CYGB-0035, an almost complete skeleton also showing soft parts, such as the flight membrane and some fur. The type species is N. liuae. The genus name refers to the Ningcheng district in Inner Mongolia; the specific name honours the collector of the fossil Ms Liu Jingyi. Ningchengopterus liuae - новый птерозавр из группы птеродактилоидов, найденный в раннемеловых отложениях формации Yixian, Внутренняя Монголия, Китай. Останки представляют собой детеныша летающего ящера, у которого сохранилась мягкая ткань, а также почти полный скелет с черепом. Этот род и вид характеризуется черепом, длина которого немного больше, чем общая длина спинных и крестцовых позвонков; 50 зубами (включая верхние и нижние челюсти); короткими средними шейными позвонками; лопаткой и плечевой костью равными в сумме по длине метакарпалю, поддерживающему крыло; локтевой костью и бедром по длине равными первой и третьей фаланге крыла соответственно. Схожие соотношения второй и первой фаланг крыла у нингченгоптеруса и Eosipterus (ёзиптерус) говорят о том, что первый систематически был близок к семейству ктенохазмид, однако это также может означать и разницу в росте отдельных фаланг крыла. Длина черепа ящера - всего 3,8 см, лопатки - 1,5 см, размах крыльев у этого "птенца" был не больше 20 см. Челюсти очень тонкие, но уже с заметными острыми зубами.    Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, птеродактили, птерозавры |
| September 27th, 2014 | |
| 04:55 pm [industrialterro] [Link] |
Nemicolopterus Nemicolopterus is a genus of pterodactyloid pterosaur, described in 2008. The type and only known species is N. crypticus. It lived in the Jehol Biota 120 million years ago. Its wingspan of slightly under 25 centimeters (10 in) makes it smaller than any but a few specimens of hatchling pterosaurs. The specimen is not fully grown, but Wang et al. (2008) cite the amount of bone fusion and the ossification of the toes, gastralia and sternum to indicate that it was a sub-adult rather than a hatchling. Darren Naish argued on his popular weblog that, due to the fact pterosaurs are highly precocial, bone fusion and ossification could occur very early, and Nemicolopterus might in fact be a hatchling individual of the genus Sinopterus. The generic name "Nemicolopterus" comes from the Greek words as follows. "Nemos" means "forest", "ikolos" means "dweller", and "pteros" means "wing". The specific name crypticus is from "kryptos", meaning "hidden". Thus "Nemicolopterus crypticus" means "Hidden flying forest dweller". N. crypticus is known from one fossil, given the catalog number IVPP V-14377. It is in the collection of the Institute of Vertebrate Paleontology and Paleoanthropology in Beijing, China. The fossil was collected from the Jiufotang Formation, which is of Aptianage (120 mya). It was discovered in the Luzhhouou locality of Yaolugou Town, Jianchang County, Huludao City, western Liaoning Province in northeastern China. Nemicolopterus is a toothless pterosaur. Wang et al. (2008) concluded that it is a primitive intermediate between the toothed pterosaurs of the Ornithocheiroidea and the often toothless pterosaurs of the Dsungaripteroidea. Though Nemicolopterus is tiny, some of the members of these groups eventually evolved into the largest flying animals that ever lived, like Quetzalcoatlus. Nemicolopterus also demonstrates clear adaptations of the toes and claws for grasping tree branches. Most pterosaurs are known from marine sediments, meaning that they probably caught fish in the ocean and landed on the adjacent beaches or cliffs. Nemicolopterus, on the other hand, is one of just a few known pterosaurs that lived in the continental interior, and probably hunted insects and roosted in the forest canopy. Its worth to note the contemporaneous pterosaur linage Tapejaridae (such as Sinopterus, which Nemicolopterus might actually be synonymous with), however, also shows strong adaptations to climbing. Самый небольшой вид древних летающих ящеров был обнаружен в Китае. Размах крыльев у найденного птерозавра-подростка составляет всего 25 сантиметров – совсем как у обычного домового воробья. Окаменелости этого крохотного ящера были найдены в провинции Ляонин. Ему было присвоено имя Nemicolopterus crypticus, что можно перевести как "скрытный летающий лесной житель". Этот маленький птерозавр представляет собой не только новый вид, но и новую группу отряда птерозавров, ранее неизвестную. А жил этот птерозавр примерно 120 миллионов лет назад. Скелет птерозавра Nemicolopterus crypticus сохранился почти полностью, что позволило сделать ряд интересных и важных выводов. Так, изученный экземпляр ещё не вполне достиг размеров и возраста взрослой особи, хотя по всем признакам вылупился из яйца давно. Попадавшиеся палеонтологам ранее, образцы птерозавров меньшего размера, были явно моложе, на момент своей гибели, данной особи птерозавра. Потому, на данный момент, это самый маленький птерозавр из всех найденных. Птерозавры жили от 228 до 65 миллионов лет назад и насчитывали, по оценкам учёных, даже больше видов, чем современные птицы. По мнению авторов работы, данное существо открывает новую главу в эволюции летающих рептилий, поскольку ранее не были известны примеры столь хорошей адаптации птерозавров к жизни в кронах деревьев. Получается, что Nemicolopterus crypticus хозяйничал среди ветвей больше, чем какой-либо другой известный птерозавр, и данное открытие расширяет не только разнообразие видов птерозавров, и без того — феноменальное, но и разнообразие мест их обитания, и соответственно - их образа жизни. Учёные выяснили, что беззубый Nemicolopterus crypticus обладает рядом анатомических особенностей, выделяющих его среди всех птерозавров. Например, необычно сильный изгиб костей стопы, показавший исследователям, что этот птерозавр-воробей значительную часть времени проводил на деревьях, обитая в пологе леса и питаясь насекомыми. Отмечен и ряд других отличий в строении черепа и конечностей. Положение птерозавра Nemicolopterus crypticus на эволюционном древе летающих ящеров указывает на его родство с более крупными птерозаврами, в том числе с видами, обладавшими размахом крыльев более 10 м, а по некоторым данным и до 18 м. Учёные считают, что группа птерозавров, к которой относится птерозавр-воробей Nemicolopterus crypticus, родственна группе Ornithocheiroidea. Учёные считают, что в целом птерозавры постепенно эволюционировали от небольших видов к крупным, научившись на каком-то этапе развития питаться рыбой, вместо насекомых, как это делал Nemicolopterus crypticus. Известно, что многие виды птерозавров обитали именно на берегу морей и океанов, где и добывали себе пропитание.
Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| 04:45 pm [industrialterro] [Link] |
Montanazhdarcho Montanazhdarcho is a genus of azhdarchid pterosaur from the late Cretaceous Period (Campanian stage) of North America, known from only one species, M. minor. The genus was named in 1993 by Kevin Padian, Armand de Ricqlès, and Jack Horner, again published by the same authors in 1995 and fully described in 2002. The type species is Montanazhdarcho minor. The genus name refers to Montana and to the related species Azhdarcho. The specific name means "the smaller one" in Latin, a reference to the relatively small size in comparison to closely related forms. The holotype, MOR 691 (Museum of the Rockies), was found by Robert W. Harmon inGlacier County, in the territory of the Blackfoot, in sandstone of the Upper Two Medicine Formation, a layer about 74 million years old. The fossil is largely uncompressed and that of an adult exemplar, as established by a study of the bone by de Ricqlès. It consists of a partial left wing, lacking the outer three wing finger phalanges, a complete shoulder girdle, a crushed cervical vertebra and two fragments of the symphysis of the mandible. The jaws were edentulous, i.e.: they lacked teeth. Montanazhdarcho was by the authors assigned to the Azhdarchidae, mainly based on the elongated form of the neck vertebra. Compared to other azhdarchids, it was small; the fragments of humerus, radius, and carpal suggest an animal with a 2.5 meter wingspan (eight feet). Its ulna was longer than the wing metacarpal, which is atypical for azhdarchids. Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархиды, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| 04:06 pm [industrialterro] [Link] |
Ludodactylus Ludodactylus was a genus of pterodactyloid pterosaurs from the Aptian-age Lower Cretaceous Crato Formation of Ceará,Brazil. The genus was named by Eberhard Frey et al. in 2003 and contains one known species, Ludodactylus sibbicki. The name is derived from Latin ludus, "play" and Greek daktylos, "finger". Ludus refers to the fact, long lamented by paleontologists, that many toy pterosaurs combined teeth with a Pteranodon-like head crest, while no such creature was known to exist — however Ludodactylus shows exactly this combination of features. "Dactylus", in reference to the characteristic long wing finger, has been a common element in the names of pterosaurs since the first known was named Pterodactylus. The specific name"sibbicki" is an homage to the paleoartist John Sibbick.
Ludodactylus is based on holotype SMNK PAL 3828, a skull missing part of the head crest, that was removed from the plate before the fossil was illegally sold. Unlike other ornithocheirids, it had no premaxillary crest on the snout, but did have a crest at the back of the skull. Frey et al. interpreted the deep mandible as a crest on the lower jaws. Trapped between the rami of the mandible is a yucca leaf; Frey suggested that the animal got it caught in its beak and unsuccessfully tried to dislodge it (the edge of the leaf is frayed), and then possibly died from starvation or a complication of starving. The skull would have been more than 66 cm (26 in) long. Frey et al. in 2003 classified Ludodactylus in the family Ornithocheiridae. In 2007 Frey, had reconsidered the validity of Ludodactylus, suggesting that it may represent the same animal as, and be a junior synonymof, Brasileodactylus. However, Andres & Myers (2013), in a large cladistic analysis of pterosaurs, found Ludodactylus to be slightly more closely related to ornithocheirids and anganguerids than to Brasileodactylus. In the analysis of Andres and Myers,Ludodactylus is classified just outside Ornithocheiridae and Anhangueridae as a derived member of the more inclusive group Pteranodontoidea.       Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| June 11th, 2014 | |
| 07:23 pm [industrialterro] [Link] |
Lonchodectes Lonchodectes (meaning "lance biter") was a genus of pterosaur from several formations dating to the Turonian (Late Cretaceous, though possibly as early as the Valanginian) of England, mostly in the area around Kent. The species belonging to it had been assigned to Ornithocheirus until David Unwin's work of the 1990s and 2000s, and the genus is not universally accepted as distinct. Several potential species are known; most are based on scrappy remains, and have gone through several other generic assignments. The genus is part of the complex taxonomy issues surrounding Early Cretaceous pterosaurs from Brazil and England, such as Amblydectes, Anhanguera, Coloborhynchus, and Ornithocheirus. Numerous species have been referred to this genus over time, and only those more widely connected with the genus are included here. The type species, L. compressirostris, is based on BMNH 39410, a partial upper jaw from the Turonian-age Upper Cretaceous Upper Chalk near Kent. Richard Owen named in 1851 as a species of Pterodactylus; it was transferred to Ornithocheirus in 1870 by Harry Govier Seeley, before becoming the type species of Lonchodectes in Reginald Walter Hooley's 1914 review of Ornithocheirus. Confusingly, this species was also long regarded, incorrectly, as the type species of Ornithocheirus. Hooley added two other species at this time, both of which had also been originally referred to Pterodactylus, then to Ornithocheirus: L. giganteus, a Cenomanian-age jaw fragment; and L. daviesii, another jaw fragment, from an Albian-age formation. The genus acquired several more former Pterodactylus and Ornithocheirus species in the 1990s-2000s. L. sagittirostris, based on BMNH R1823, a lower jaw fragment from the ?Valanginian-Hauterivian-age Lower Cretaceous Hastings Beds of East Sussex, and L. platystomus had been acquired by 2000. Two additional species based on jaw fragments, both from the Albian-age Cambridge Greensand, were added by 2003: L. machaerorhynchus and L. microdon, joining L. compressirostris, L. giganteus, L. platystomus, and L. sagittirostris in his listing of valid species. However, L. giganteus, L. machaerorhynchus, and L. microdon have since been assigned to a new genus, Lonchodraco, while L. sagittirostris has been removed from Lonchodectes and treated as a nomen dubium at Pterodactyloidea incertae sedis. The genus is poorly known and not universally accepted, as noted above. In Peter Wellnhofer's 1991 The Illustrated Encyclopedia of Pterosaurs the other major recent synopsis of pterosaurs, written before Unwin's work, the species are included with Ornithocheirus (because of L. compressirostris being thought to be the type species), and are in fact the main fossils illustrated to represent the genus. Unwin placed them in their own family, Lonchodectidae, which he grouped with the ctenochasmatoids in 2003, and with the azhdarchoids, including the tapejarids and azhdarchids, in 2006. Unwin considers Lonchodectes to have been a generalist, like a seagull, with its conservative jaws and teeth (like those of the much older Pterodactylus) and small to medium size; he estimates its maximum wingspan at about 2 m (6.6 ft). Lonchodectes had long jaws with many short teeth, and the jaws were compressed vertically, like "a pair of sugar tongs with teeth". At least some of these species had crests on their lower jaws. Recent studies, however, show that it had limb proportions akin to those of azhdarchids, and might have lived similarly. Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| June 7th, 2014 | |
| 02:53 pm [industrialterro] [Link] |
Liaoningopterus Liaoningopterus, sometimes misspelled "Liaoningopteryx", was a genus of ornithocheirid pterodactyloid pterosaur from the Barremian-Aptian-age Lower Cretaceous Jiufotang Formation of Chaoyang, Liaoning, China. The genus was named in 2003 by Wang Xiaolin and Zhou Zhimin. The type species is Liaoningopterus gui. The genus name is derived from Liaoning and a Latinised Greek pteron, "wing". The specific name honours Professor Gu Zhiwei, an invertebrate specialist who has pioneered the study of the Jehol Biota. The genus is based on holotype IVPP V-13291, a crushed partial skull and skeleton including the jaws, teeth, a cervical vertebra, and bones of the finger supporting the wing. It was a large pterosaur — the largest known from China at the time of description — with a skull length estimated at 61 cm (24 in), and a wingspan estimated at five metres (16.4 ft). The skull was long and low, bearing low crests close to the tip of the beak on both lower and upper jaws. The snout crest was twelve centimetres long, was symmetrical in form and had a maximum height of seventeen millimetres. The edge of the upper jaw was very straight. The teeth were only found at the anterior end of the jaws. They were elongated but robust, generally increasing in size from the back to the front. The fourth tooth in the upper jaw is with a length of 81 millimetres the largest known for any pterosaur. It is exceptional in size compared to the other teeth of Liaoningopterus also, the longest tooth in the lower jaw having a length of 41 millimetres. Tooth length in the specimen is very variable, which the authors explained by the presence of recently erupted replacement teeth. There were twenty pairs of teeth in the upper jaws and thirteen or fourteen pairs in the lower jaws. The preserved cervical vertebra has a centrum length of 46 millimetres and a centrum height of 34 millimetres. From the wing bones pieces of the first phalanx can be recognised which had an estimated total length of about fifty centimetres. The authors described Liaoningopterus as being probably a piscivore, due to the long, pointed snout. Wang classified Liaoningopterus as a member of the Anhangueridae, mainly because of the crests. This opinion was restated by him in 2005. In 2006 Lü Junchang published a cladistic analysis showing Liaoningopterus to be a basal member of the Anhangueridae; in 2008 an analysis by Ji Qiang had Liaoningopterus in a trichotomy with Anhanguera and Tropeognathus. Tags: Вымершие рептилии, Мел, авеметатарзалии, аньянгуэриды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| 01:32 pm [industrialterro] [Link] |
Lacusovagus Лакусовагус (Lacusovagus; «озерный странник») — род птерозавров семейства Chaoyangopteridae, обитавший в меловом периоде примерно 115 млн лет назад. Ископаемые останки, найденные в отложениях на северо-востоке Бразилии, были описаны в 2008 году. Сохранились только фрагменты черепа. Размах крыльев составлял около пяти метров. Lacusovagus (meaning "lake wanderer") is a genus of azhdarchoid pterodactyloid pterosaur from the Lower Cretaceous of Brazil. It is based on SMNK PAL 4325, a partial upper jaw comprising sections of the skull in front of the eyes. This specimen was found in rocks of the Early Cretaceous-age (probably Aptian stage, about 120 million years ago) Nova Olinda Member of the Crato Formation. The skull was long, and unusually wide. The section in front of the combined nasal-antorbital fenestra was relatively short. Also unusual was the combination of its toothless jaws and no bony head crest. Lacusovagus was described in 2008 by Mark Witton. The type species is L. magnificens, meaning "grand lake wanderer", in reference to its large size — it is currently the largest pterosaur known from the Crato Formation with an estimated wingspan of 4.1 meters (13 feet). Lacusovagus shares many characteristics with the basal azhdarchoid family Chaoyangopteridae, and preliminary studies suggested it was a member of that clade. The Chaoyangopteridae are a family of pterosaurs within the Azhdarchoidea. The clade Chaoyangopteridae was first defined in 2008 by Lü Junchang and David Unwin as: "Chaoyangopterus, Shenzhoupterus, their most recent common ancestor and all taxa more closely related to this clade than to Tapejara, Tupuxuara or Quetzalcoatlus". Based on neck and limb proportions, it has been suggested they occupied a similar ecological niche to that of azhdarchid pterosaurs, though it is possible they were more specialised as several genera occur in Liaoning, while azhdarchids usually occur by one genus in a specific location. The Chaoyangopteridae are mostly known from Asia, though the possible member Lacusovagus occurs in South America and there are possible fossil remains from Africa. Учёные определили новый вид птерозавра. Это самый крупный хаоянгоптерид из всех известных и единственный в своёй группе, найденный за пределами Китая. С классификацией огромной твари разобрался Марк Уиттон (Mark Witton) из университета Портсмута (University of Portsmouth). «Ранее в Китае уже находили останки животных этого семейства (Chaoyangopteridae), — рассказывает Марк Уиттон. — По величине они не превышали 60 сантиметров, что сравнимо с размерами черепа обнаруженного в Бразилии экземпляра. Размах его крыльев должен был составлять около пяти метров». Исследователь дал новому виду имя Lacusovagus («озерный странник»). При идентификации образца у исследователя возникли проблемы: обычно останки из этих районов Бразилии поступают в прекрасном (по меркам палеонтологии) состоянии, однако череп Lacusovagus был сильно сдавлен. Даже то, что у рептилии не было зубов, ученому пришлось доказывать косвенно. С уверенностью можно говорить лишь о том, что череп птерозавра был очень широк; возможно, это позволяло ему охотиться на весьма крупных животных. «Впрочем, — заключает Марк Уиттон, — пока мы располагаем лишь отрывочными данными. Необходимо отыскать еще несколько образцов — вот тогда можно будет сделать определенные выводы». По заверениям исследователей, обитал этот ящер на больших озёрах, за что и получил латинское обозначение вида . Стоящий на земле птерозавр превышал метр в плечах (а подняв голову, оказывался бы сравним по росту со взрослым человеком). Ну а расправив крылья, это чудище могло похвастать пятиметровым их размахом. Любопытно, что перед отправкой на анализ к Марку и его коллегам этот экземпляр птерозавра пролежал в одном из немецких музеев несколько лет. Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры, хаоянгоптериды |
| June 4th, 2014 | |
| 07:46 pm [industrialterro] [Link] |
Istiodactylus Istiodactylus is a genus of a medium sized pterosaurs from the early Cretaceous period. Istiodactylus were moderately large pterosaurs. Correcting earlier estimates of a length of fifty-six centimetres, Mark Paul Witton in 2012 concluded that the skull of one specimen, NHMUK R3877, was forty-three centimetres long. The maximum wingspan was probably about 4.3 metres (14.1 ft), making the largest individuals roughly half the size of the largest Pteranodon. Because of the flat, rounded shape of the snout, Istiodactylus is sometimes referred to as a "duck-billed pterosaur". However, unlike ducks, they had teeth. These triangular teeth were laterally compressed, an adaptation for slicing flesh, and interlocked tightly in the closed jaws. The skull was relatively short, with most teeth concentrated in the blunt front tip of the jaws. Witton's 2012 study proposed that Istiodactylus were primarily scavengers. In 1887 Harry Govier Seeley named the remains of a fossil pelvis discovered on the Isle of Wight, Ornithodesmus cluniculus, thinking it was a bird. In 1901 he considered that it might have been a pterosaur. In 1913 Reginald Walter Hooley named a second species, Ornithodesmus latidens, for some definitively pterosaurian material, found in the Vectis Formation. The holotype was BMNH R 0176, a partial skeleton. The specific name means "broad tooth" in Latin. The type species of the genus Ornithodesmus, however, was in the 1980s discovered to be based on bones belonging to a dinosaur, which meant a new genus had to be named for O. latidens. This species was assigned to its own genus Istiodactylus by Stafford Howse, Andrew Milner, and David Martill in 2001. The genus name is derived from Greek istion, "sail" and daktylos, "finger", referring to the fact that the wing of pterosaurs is formed by a membrane attached to a wingfinger. Howse et al. in 2001 created for Istiodactylus its own family Istiodactylidae. In 2006 a second species, I. sinensis from China, has been named, its specific name referring to China. Its holotype is NGMC 99-07-011, a partial skeleton of a subadult individual. It was much smaller than I. latidens, its dimensions being 63% of the larger species, thus about a quarter in weight. In 2006 Lü Jun-Chang et al. concluded I. sinensis was a junior synonym of the istiodactylid Nurhachius. Mark Witton has proposed it may be a synonym of Liaoxipterus. Istiodactylidae is a small family of pterosaurs. This family was named in 2001 after the type genus Istiodactylus was discovered not to be a member of the genus Ornithodesmus. Remains of taxa that can be confidently assigned to Istiodactylidae have been found in the UK and China, in rocks dating from the Early Cretaceous period (Barremian to Aptian stage). Arbour and Currie (2011) described Canadian Gwawinapterus beardi as a member of Istiodactylidae living in the late Cretaceous (upper Campanian stage); however, Witton (2012) suggested the tooth replacement pattern in this animal does not match that of pterosaurs, suggesting that the species might be non-pterosaurian. Additional research suggested that the species was in fact a fish. The earliest known species might be Archaeoistiodactylus linglongtaensis, from the Middle Jurassic of China; however, it also has been suggested that the holotype specimen of this species might actually be a poorly preserved specimen of Darwinopterus. Hongshanopterus, a supposed istiodactylid from China, has been reclassified as a non-istiodactylid member of Ornithocheroidea of uncertain phylogenetic placement by Witton (2012). Istiodactylids were medium sized pterosaurs with flat, rounded jaws similar to that of a duck. They had small teeth lining their jaws, however. Крылатые рептилии мелового периода были не хищниками, а самыми настоящими падальщиками наподобие современных грифов. Питались они главным образом мертвыми динозаврами. Неожиданное открытие сделал доктор Марк Уиттон из Портсмутского университета. Он решил тщательно изучить останки птерозавра Istiodactylus latidens, найденного в 1904 году на острове Уайт, и с тех пор лежавшего на витрине Лондонского музея естественной истории. Поскольку до сих пор изученными оставались лишь черепная коробка и ветви челюсти этого животного, у палеонтологов сложилось мнение о его узкой, низкой и длинной голове. Как показало новое исследование, голова истиодактилуса, напротив, была высокой, широкой и короткой. При этом кости обеспечивали надежное крепление чудовищной шейной мускулатуре, позволявщей крылатому ящеру легко отрывать куски плоти с туш крупных животных. В то время как зубы Istiodactylus latidens были острыми и приспособленными для разрезания мяса, его скулы оказались неожиданно тонкими. Их толщина едва достигает шести миллиметров при общей высоте черепа до 45 сантиметров. Ящер легко мог сломать себе кости, если бы вступил в схватку даже с не слишком сильным противником, сообщает The Daily Mail. Именно это обстоятельство позволило доктору Уиттону сделать предположение о том, что добыча истиодактилусов никогда не сопротивлялась, а просто лежала на одном месте. "Современные птицы-падальщики имеют схожую конструкцию черепа, обладающую как
сильными, так и слабыми компонентами. Поедание мертвой плоти позволяет птицам-падальщикам
практически полностью контролировать процесс питания, поэтому они могут
позволить себе иметь несколько слабых участков, расположенных вдоль челюсти, не
беспокоясь о том, что их можно сломать при укусе, – рассказал палеонтолог. –
Широкий и высокий череп Istiodactylus latidens позволяет прикрепить
мощный набор шейных мышц, что указывает на сильную шею, которая является еще
одним общим качеством птиц-падальщиков, которые используют свои сильные шеи, чтобы
растаскивать части трупа. Если сопоставить данные обстоятельства с режущими
зубами, то все эти свойства являются хорошим доказательством падалеядного образа жизни, который вели эти животные". Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, истиодактилиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| May 30th, 2014 | |
| 07:38 pm [industrialterro] [Link] |
Hatzegopteryx Hatzegopteryx (рум. Hațeg — название румынского города, πτέρυξ — крыло) — род аждархидовых птерозавров, известный по останкам, найденным в Трансильвании в 2002 году. Черепные фрагменты, левая плечевая кость, и другие ископаемые останки указывают на то, что Hatzegopteryx был одним из самых крупных птерозавров. Род был назван в 2002 году французским палеонтологом Eric Buffetaut и румынскими палеонтологами Dan Grigorescu и Zoltan Csiki. Включает в себя только один вид Hatzegopteryx thambema. Название рода образовано от румынского города Хацег (кости Hatzegopteryx были найдены в районе так называемого «Hațeg Island (англ.)») и греч. πτέρυξ «крыло». Видовое название thambema образовано от греч. "монстр" и отсылает к размерам Hatzegopteryx. Останки Hatzegopteryx относятся к Densus Ciula Formation в западной Румынии, которая датируется поздним маастрихтом (верхний мел, приблизительно 65 млн лет назад). Голотип, FGGUB R 1083A, состоит из задней части черепа и поврежденной проксимальной части левой плечевой кости. 38,5-сантиметровый диафиз бедренной кости, найденный поблизости (FGGUB R1625), может тоже принадлежать Hatzegopteryx. Hatzegopteryx, предположительно, имел крепкий расширяющийся в задней части череп и массивные челюсти. Многие ископаемые кости Hatzegopteryx сильно схожи с костями Quetzalcoatlus sp., но у Hatzegopteryx был намного более тяжелый череп и другое сочленение челюстей, похожее на таковое у птеранодона. Сравнивая с черепами других птерозавров: Nyctosaurus и Anhanguera, Buffetaut с коллегами, при описании найденных образцов дают оценку длины черепа Hatzegopteryx до 3 м, что больше, чем у кетцалькоатля. Авторы оценивали размер Hatzegopteryx, сравнивая фрагмент плечевой кости длиной 23,6 см, с плечевой костью кетцалькоатля, считавшегося ранее самым крупным известным птерозавром. Образец кетцалькоатля TMM 41450-3 имеет плечевую кость длиной 54,4 см. Фрагмент плечевой кости Hatzegopteryx меньше половины целой, поэтому она, скорее всего, была немного длиннее кости кетцалькоатля. Размах крыльев последнего был в 1981 году оценен в 11—12 метров, более ранние оценки же говорили о 15—20 метрах. Отсюда авторы заключили, что оценка размаха крыла в 12 метров достаточно скромна, «если его плечевая кость была действительно длиннее, чем таковая у Q. northropi». В 2003 году оценки были пересмотрены: размах крыльев Hatzegopteryx был оценен приблизительно в 12 метров, а длина черепа — свыше 2,5 метра. В 2010 году Mark Witton заявил, что кажущееся превосходство размеров плечевой кости Hatzegopteryx вызвано её деформацией после отложения в породе, так что, вероятно, размах крыльев Hatzegopteryx был не больше, чем размах крыльев кетцокоатля, сегодня обычно оцениваемый в 10—11 метров. ( Read More ) Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9): ( Read More ) Размеры тела в сравнении с человеком и жирафом: Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархиды, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| May 28th, 2014 | |
| 05:48 pm [industrialterro] [Link] |
Guidraco Guidraco (Chin. gui (鬼) "malicious ghost" + Lat. draco "dragon") is an extinct genus of toothed pterodactyloid pterosaur known from the Early Cretaceous of Liaoning Province, northeast China. Guidraco is known only from the holotype IVPP V17083, an articulated partial skeleton consisting of a nearly complete skull, lower jaws and a series of four, second to fifth, cervical vertebrae. It was collected at Sihedang near Lingyuan City in the Liaoning Province from the Jiufotang Formation, dating to the Aptian stage of the Early Cretaceous, about 120 million years ago. Guidraco was first described and named by Wang Xiaolin, Alexander W.A. Kellner, Jiang Shunxing and Cheng Xin in 2012 and the type species is Guidraco venator. The generic name is derived from Chinese gui (鬼), "malicious ghost", and from Latin draco, "dragon". The specific name means "hunter" in Latin. The holotype skull of Guidraco has a length of thirty-eight centimetres. It is very elongated with a hollow profile but not very pointed as the upper edge and the line of the jaw run nearly parallel over most of their length. The ensuing relative robustness of the snout is reinforced by a short main skull opening, a fenestra nasoantorbitalis with just a quarter of skull length, and a lower jaw equalling the rostrum in depth. The snout lacks a crest. Above the eye sockets however, the line of the top of the snout curves steeply upwards, resulting in a very large crest on the frontals, as high as the posterior part of the skull is deep, ending in a rounded top. Due to the angling of the skull roof the crest slightly points forwards and its base extends to the back of the roof; however, the parietal is not part of it. In front of the crest large impressions of soft tissue are visible but these are plant remains. Further diagnostic features of the skull include an infratemporal fenestra with a narrow lower end, and a jugal of which the front branch extends no further than the front edge of the fenestra nasoantorbitalis. The teeth of Guidraco are very distinctive. Of the twenty-three teeth of the upper jaw the first is long and very narrow, pointing nearly horizontally forward. The next three teeth are enormous in size, very long, robust, pointed and slightly recurved. They gradually point more downwards. These are followed by a series of three medium-length downward-pointing straight teeth, of which the middle one, the sixth, is the shortest. The remaining thirteen teeth constitute a long row of small elements gradually diminishing in size. This arrangement is mirrored by the eighteen teeth of the lower jaw. Here however, a forward pointing tooth is lacking. The first four teeth are of great size, even longer than their counterparts of the upper jaw. Next is a series of three straight teeth of medium height, followed by a row of eleven increasingly smaller elements for a grand total in the head of eighty-two teeth. With the fossil, the beak is closed and due to their extreme length the front teeth extend far beyond the upper and lower edges of the head, the protruding parts being up to twice as long as the depth of the snout or lower jaw. The teeth can also be divided into two types according to their built: the first nine teeth of the upper jaw and eight teeth of the lower jaw have vertical ridges on the back of their enamel; the back teeth have a uniformly smooth enamel and thickened crown bases, giving them a more triangular outline. Though not having the form of a true rosette because the jaw ends were not expanded, the intermeshing front teeth functioned as a "prey grab" to catch slippery animals; the describers therefore consider Guidraco to have been a fish-eater. The neck vertebrae are moderately elongated, keeled and possess large pneumatic openings on their sides, the access by which the air sac of the neck could enter their hollow interiors. The axis bears a spiked spine. Guidraco was by the describers assigned to the Pteranodontoidea sensu Kellner. A phylogenetic analysis found it to be the sister taxon of the Brazilian Ludodactylus, the two species together forming a clade that was closely related to the Istiodactylidae and the Anhangueridae. The fact that a Chinese form is closely related to a South-American species would indicate a large faunal interchange between continents in this period. Китайская академия наук на днях опубликовала документы, согласно которым на северо-востоке страны ученые обнаружили останки птерозавра редкого типа, имевшего длинные зубы устрашающего вида. Согласно опубликованным фотографиям, доисторическая летающая рептилия имела мощные челюсти и крупные резцовые зубы. Палеонтологи говорят, что найденный вид является довольно близким биологическим родственником вида Ludodactylus, останки которого в прошлом были обнаружены учеными на территории современной Бразилии. Согласно данным исследователей, рептилия обитала на нашей планете в ранний меловой период - как раз в расцвет эпохи динозавров. Останки ящера были найдены на территории современной китайской провинции Ляонин, где ранее уже находили останки летающих ящеров. Ископаемые останки и реплики (1, 2, 3, 4): Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, бореоптериды, диапсиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| 04:03 pm [industrialterro] [Link] |
Geosternbergia Geosternbergia from the Late Cretaceous geological period of North America, was one of the largest pterosaur genera and had a wingspan of up to 7.25 metres (23.8 ft). Geosternbergia was among the largest pterosaurs, with the wingspan of most adults ranging between 3 and 6 meters (9.8–19.6 ft). While most specimens are found crushed, enough fossils exist to put together a detailed description of the animal. Geosternbergia is distinguished from Pteranodon mainly by its upright-crest. The lower jaw of G. sternbergi was 1.25 meters (4 ft) long. The most distinctive characteristic of Geosternbergia is its cranial crest. These crests consisted of skull bones (frontals) projecting upward and backward from the skull. The size and shape of these crests varied due to a number of factors, including age, gender, and species. Male G. sternbergi, the older species of the two described to date, had a larger vertical crest with a broad forward projection, while G. maysei had a short, rounded vertical crest and was generally smaller. Females of both species were smaller and bore small, rounded crests. The crests were probably mainly display structures, though they may have had other functions as well. It was collected by George F. Sternberg in 1952 and described by John Christian Harksen in 1966, from the lower portion of the Niobrara Formation. It was older than Pteranodon longiceps and is considered by Bennett to be the direct ancestor of the later species. Geosternbergia fossils are known from the Niobrara and Sharon Springs Formations of the central United States. Geosternbergia existed as a group for more than four million years during the late Coniacian - early Campanian stages of the Cretaceous period. The genus is present in the lower the Niobrarra Formation except for the upper two; in 2003, Kenneth Carpenter surveyed the distribution and dating of fossils in this formation, demonstrating that Geosternbergia sternbergi existed there from 88-85 million years ago, while the species later named G. maysei existed between 81.5 and 80.5 million years ago. Geosternbergia was traditionally considered a species, or occasionally subgenus, of the similar pterosaur Pteranodon, in most major studies of pteranodontians through the 1990s. However, a 2010 review of the group by A.W.A. Kellner suggested that Pteranodon sternbergi was different enough from P. longiceps to belong in a distinct genus, to which Kellner also referred a new species, Geosternbergia maysei. Earlier, pterosaur researcher Chris Bennett had considered the G. maysei specimen an adult male P. longiceps. Adult Geosternbergia specimens may be divided into two distinct size classes, small and large, with the large size class being about one and a half times larger than the small, and the small being twice as common as the large. Both size classes lived alongside each other, and while researchers had previously suggested that they represent different species, Christopher Bennett showed that the differences between them are consistent with the concept that they represent females and males, and that Geosternbergia species were sexually dimorphic. Skulls from the larger size class preserve large, upward and backward pointing crests, while the crests of the smaller size class are small and triangular. Some larger skulls also show evidence of a second crest that extended long and low, toward the tip of the beak, which is not seen in smaller specimens. The gender of the different size classes was determined, not from the skulls, but from the pelvic bones. Contrary to what may be expected, the smaller size class had disproportionately large and wide-set pelvic bones. Bennett interpreted this as indicating a more spacious birth canal, through which eggs would pass. He concluded that the small size class with small, triangular crests represent females, and the larger, large-crested specimens represent males. Note that the overall size and crest size also corresponds to age. Immature specimens are known from both females and males, and immature males often have small crests similar to adult females. Therefore, it seems that the large crests only developed in males when they reached their large, adult size, making the gender of immature specimens difficult to establish from partial remains. The fact that females appear to have outnumbered males two to one suggests that, as with modern animals with size-related sexual dimorphism, such as sea lions and other pinnipeds, Geosternbergia might have been polygynous, with a few males competing for association with groups consisting of large numbers of females. Similar to modern pinnipeds, Geosternbergia may have competed to establish territory on rocky, offshore rookeries, with the largest, and largest-crested, males gaining the most territory and having more success mating with females. The crests of male Geosternbergia would not have been used in competition, but rather as "visual dominance-rank symbols", with display rituals taking the place of physical competition with other males. If this hypothesis is correct, it also is likely that male Geosternbergia played little to no part in rearing the young; such a behavior is not found in the males of modern polygynous animals who father many offspring at the same time. The Pteranodontidae are a family of large pterosaurs of the Cretaceous Period of North America. The family was named in 1876 by Othniel Charles Marsh. Pteranodontids had a distinctive, elongated crest jutting from the rear of the head (most famously seen in Pteranodon itself). The spectacularly-crested Nyctosaurus is sometimes included in this family, though usually placed in its own family, the Nyctosauridae (Nicholson & Lydekker, 1889). Modern researchers differ in their use of the concept. S. Christopher Bennett and Alexander Kellner have concluded that Nyctosaurus was not a pteranodontid. In 1994 Bennett defined a clade Pteranodontidae, also including species of the Anhangueridae. However, this definition has not been accepted by other workers. Alexander Kellner, for example, named several additional species for specimens previously classified as Pteranodon, and placed P. sternbergi in a distinct genus, Geosternbergia. Kellner re-defined Pteranodontidae as the most recent common ancestor of Pteranodon longiceps, Geosternbergia sternbergi and Dawndraco kanzai, and all of its descendants. This clade possibly includes the nyctosaurids. Analyses by David Unwin did indicate a close relationship between Pteranodon and Nyctosaurus, and he used the name Pteranodontia for the clade containing both. Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10): ( Read More ) Размеры тела в сравнении с человеком для самцов и самок: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеранодонтиды, птеранодонтойды, птеродактили, птерозавры |
















































































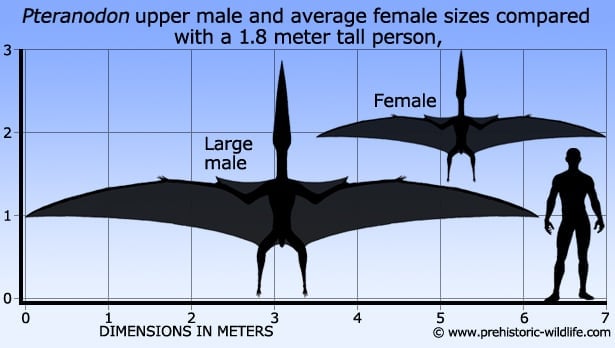



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}