[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 20 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| March 19th, 2012 | |
|---|---|
| 07:38 pm [industrialterro] [Link] |
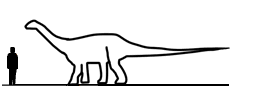
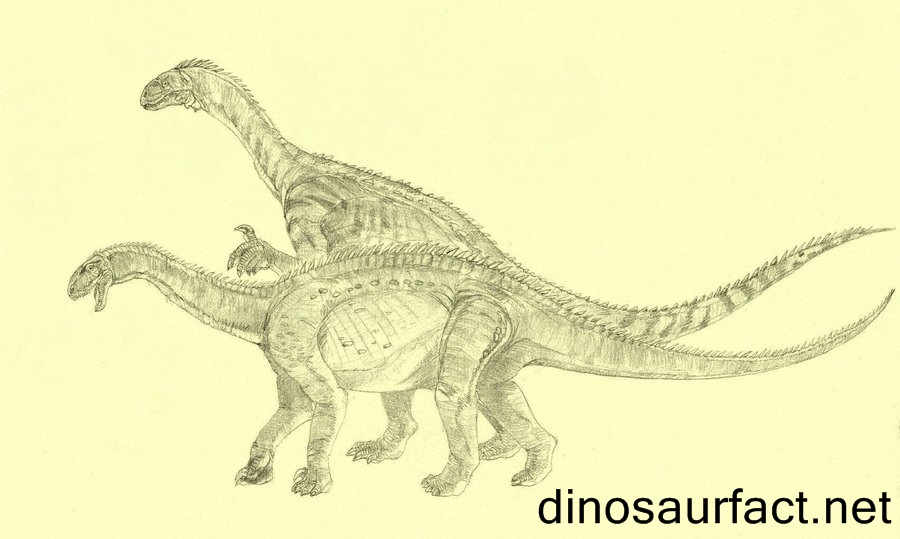
Plateosaurus Платеоза́вр (Plateosaurus — «плоский ящер») — крупнейший из известных динозавров триасового периода. Его длина достигала 6—10 м, масса — до 700 кг. Как и все завроподоморфы, платеозавр принадлежит к ящеротазовым. Впервые описан немецким палеонтологом Германом фон Майером в 1837 году, который и придумал ему имя. Жил на территории современной Европы в верхнем триасе 220 - 200 млн. лет назад. Будучи травоядным прозауроподом, платеозавр имел длинную шею и приземистое туловище грушевидной формы. Череп — маленький и узкий по сравнению с размерами туловища. Зубы — ланцетовидные (в верхней челюсти свыше 30, в нижней — менее 30). Морда — вытянутая, глаза направлены в стороны, а не вперед, что улучшало обзор и позволяло заранее заметить хищника. Передние конечности были короче задних и на них четко выделялись пальцы. Это свидетельствует о том, что они были приспособлены для хватания, то есть могли использоваться для хватания пищи. Среда естественного обитания платеозавров — пустынные территории Европы. Его окаменелости наиболее типичны для Европы, где найдено всего более 50 мест его обитания. Платеозавр был крупнейшим динозавром триасового периода. Их размеры и масса залог их успеха. В триасовый период у динозавра весом в 4 тонны вряд ли нашлись бы враги. Голова платеозавра по сравнению с телом была маленькой. В пасти было около 60 ланцетовидных зубов - примерно по 30 на верхней и нижней челюсти. Череп вытянутый, глаза направлены в стороны. Челюсти слабые, приспособленные для обрывания растительности. Это улучшало обзор и помогало заметить хищников. Платеозавр перемещался на четырех лапах. Задние лапы были гораздо больше передних. Это позволяло платеозавру легко вставать на задние лапы, чтобы достать листву с высоких ветвей. Длинный хвост платеозавра служил для удержания равновесия, а также был средством защиты. Передние конечности были короче задних и при этом были хорошо развиты. Оканчивались ярко выраженными пальцами, отлично приспособленными для захвата веток. Платеозавры были стадными динозаврами. Они обитали по берегам водоемов триасового периода, или кочевали по триасовым пустыням в поисках пищи и живительной влаги. Учитывая размеры, платеозавр ежедневно нуждался в большом количестве пищи. Платеозавр, как и другие динозавры триасового и юрского периода, не пережёвывал пищу, а заглатывал вместо этого камни, с помощью которых растительная масса окончательно перетиралась и переваривалась в желудке. Пережевывать пищу приспособились только динозавры мелового периода.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8, 9, 10):
( Далее ) Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, платеозавриды, прозавроподы, ящеротазовые |
| 07:20 pm [industrialterro] [Link] |
Mussaurus Мусза́вр (Mussaurus — «мышиный ящер») — травоядный динозавр, принадлежащий к группе прозавропод и живший в позднетриасовом периоде на территории современной Аргентины. Все известные останки мусзавра принадлежат детёнышам. Скелеты их столь малы, что легко помещаются на ладони. Размеры взрослых особей неизвестны, но, вероятно, они могли достигать 5—6 метров в длину и весить до 150 кг. Первые найденные учеными семь скелетов мусзавра имели череп всего 3 сантиметра; все они находились рядом. Находка походила на остатки гнезда, а в нем лежали хорошо сохранившиеся кости маленьких мусзавров. Кто бы мог предположить, что детёныш динозавра может быть настолько мал, что мог бы уместиться на ладони! Почему же детеныши погибли? Возможно, пострадали от болезни, или погибла мать; или на них напал хищник...
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, прозавроподы, ящеротазовые |
| 07:03 pm [industrialterro] [Link] |
Melanorosaurus Melanorosaurus (meaning "Black Mountain Lizard", from the Greek melas/μέλας, "black", oros/ὄρος, "mountain" + sauros/σαῦρος, "lizard"), is a genus of basal sauropodomorph dinosaur that lived during the Late Triassic period. A herbivore from South Africa, it had a large body and sturdy limbs, suggesting it moved about on all fours. Limb bones were massive and weighty, like sauropod limb bones. The type specimens, syntypes SAM 3449 and SAM 3450, were described and named in 1924 by Sidney Henry Haughton. They were collected from the Triassic Lower Elliot Formation, dating to the early Norian, on the north slope of the Thaba 'Nyama (Black Mountain) in Transkei, South Africa. The first complete skull of Melanorosaurus was described in 2007. Two species are known: M. readi, the type species, and M. thabanensis named in 1993 by François-Xavier Gauffre, based on holotype MNHN LES-16, a femur found in the Jurassic Upper Elliot Formation. Melanorosaurus was once classified as a prosauropod, but Prosauropoda no longer appears to be a natural group. According to some definitions of Sauropoda, Melanorosaurus is an early sauropod. However, these definitions also take in many other former "prosauropods," and Adam Yates has proposed a definition of Sauropoda that will specifically exclude Melanorosaurus (Sauropoda as all sauropodomorphs closer to Saltasaurus than Melanorosaurus). This definition would allow Sauropoda to retain its traditional concept. Melanorosaurus had a skull which measured approximately 250 mm. The snout was somewhat pointed, and the skull was somewhat triangular when seen from above or below. The premaxilla had four teeth on each side, a characteristic of primitive sauropodomorphs. The maxilla had 19 teeth on each side of the jaw. Melanorosaurus was around 8 metres (26 ft) long, with a weight of 1.3 metric tons (1.3 long tons).
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, меланорозавриды, прозавроподы, ящеротазовые |
| 06:44 pm [industrialterro] [Link] |
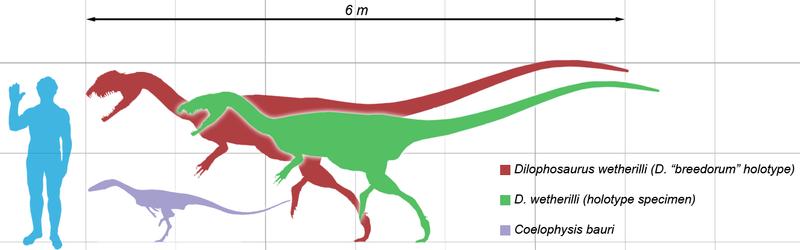
Liliensternus Liliensternus (meaning "Rühle von Lilienstern's one") was a genus of coelophysoid dinosaur from the Late Triassic period, about 205 Ma. Liliensternus was originally named in 1934 by Friedrich von Huene as a second species of Halticosaurus, H. liliensterni, the specific name honouring the German amateur paleontologist, Dr. Hugo Rühle von Lilienstern, who in 1922 and 1923 had found the remains near Großen Gleichberg in Germany's Trossingen Formation (late Norian). The naming was occasioned by the opening on 1 July 1934 of a paleontological museum at Count Rühle von Lilienstern's castle at Bedheim, where the fossils remained until 1969 when they were transferred to the Humboldt Museum in Berlin. In 1984 Samuel Paul Welles concluded that the type species of Halticosaurus, H. longotarsus, was a nomen dubium. Most what had been written in the literature about Halticosaurus in fact regarded H. liliensterni. Welles therefore named a separate genus: Liliensternus, the name again honouring Rühle von Lilienstern. The type species is Liliensternus liliensterni. A second species named in 1993 for remains found in France, Liliensternus airelensis, which had an extra pair of cervical pleurocoels, has in 2007 been renamed a separate genus, Lophostropheus. The remains, together forming a syntype series with inventory number MB.R.2175, consist of the partial and fragmentary skeletons of at least two individuals, containing elements of the skull, the lower jaws, the vertebrae and the appendicular skeleton. Liliensternus was approximately 5.15 m (17 ft) long, and may have weighed about 127 kg (280 lb). It might have preyed on herbivores like Plateosaurus. Originally assigned to the Halticosauridae, Liliensternus is today considered a basal member of the Coelophysoidea.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, целофизиды, ящеротазовые |
| 06:41 pm [industrialterro] [Link] |
Lessemsaurus Lessemsaurus is an extinct genus of sauropodomorph dinosaur (possibly a melanosaurid) named for the writer of popular science books Don Lessem. The type species, L. sauropoides, was formally described by José Fernando Bonaparte in 1999. This dinosaur was around 30 feet long, and was discovered in strata dating to the Norian stage, around 210 million years ago. The Melanorosauridae were a family of sauropodomorph dinosaurs which lived during the Late Triassic and Early Jurassic. The name Melanorosauridae was first coined by Friedrich von Huene in 1929. Huene assigned several families of dinosaurs to the infraorder Prosauropoda: the Anchisauridae, the Plateosauridae, the Thecodontosauridae, and the Melanorosauridae. Since then, these families have undergone numerous revisions. Galton and Upchurch (2004) considered Camelotia, Lessemsaurus, and Melanorosaurus members of the family. A more recent study by Yates (2007) indicates that the melanorosaurs were instead early sauropods. The two members of the Riojasauridae, Riojasaurus and Eucnemesaurus, have also been placed in Melanorosauridae, which would render the Riojasauridae an invalid taxon. Прозавропо́ды (Prosauropoda) — одна из подгрупп группы ящеротазовых динозавров. У них был длинный хвост и бочкообразное туловище. Некоторые прозавроподы передвигались на четырёх лапах, другие — только на двух. Питались они растениями и были первыми крупными динозаврами, появившимися на Земле. Жили в позднетриасовом и раннеюрском периодах (220—180 млн. лет назад). В течение триасового периода прозавроподы жили по всей Пангее, то есть по всей поверхности планеты (прим. - cуши, естественно). Несмотря на свое название и ряд общих черт, роднивших их с завроподами, эти динозавры вовсе не являлись их прародителями. Вероятно, они представляли собой побочную ветвь группы завропод. Небольшие размеры позволяли прозавроподам ходить на двух лапах. Но это они делали далеко не всегда, поскольку могли благополучно передвигаться и на всех четырёх. Поднимаясь на задние лапы, они поедали листву с верхних веток. Кроме растительной пищи прозавроподы питались и животной. Остатки прозавропод известны ещё с XIX века. Например, скелет анхизавра нашли в 1818 году, но лишь в 1885 году выяснилось, что кости принадлежат пресмыкающемуся. Остается спорным следует или нет рассматривать данную группу как парафилетическую (то есть включает лишь часть потомков гипотетического общего предка) среди базальных зауроподоморфов, или можно выделить прозавроподов как монофилетическую группу, причем в последнем случае динозавры вряд ли могут быть включены в эту группу. Galton и Upchurch (2004) определяет кладу прозауроподы, как группу, включающую платеозавра и все таксоны более тесно связанные с ним, а также все виды базальных завроподоморфов, не принадлежащих к зауроподам. Другие исследователи (например, Пол Серено, 2005) считают, однако, что прозавроподы, охватывающие все эти виды, будут являться парафилетической группой, к которой будут относится предки зауроподов, но не сами зауроподы. В последние годы некоторые ученые сделали попытку пересмотреть систематику прозавроподов, таким образом, чтобы исключить их из числа предков зауроподов. Например, Йетс и Китчинг (2003) исключили из их числа базальных прозавроподов Thecodontosaurus и Saturnalia, а также рассматриваемых ими в качестве базальных зауроподов - Anchisaurus и Melanorosaurus. Ятс (2007) пошел еще дальше, заявив, что прозавроподы, такие как Massospondylus и Riojasaurus, на самом деле, более тесно связаны с завроподами, чем с другими прозавроподами.
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, меланорозавриды, прозавроподы, ящеротазовые |
| March 18th, 2012 | |
| 05:48 pm [industrialterro] [Link] |
Isanosaurus Isanosaurus (meaning "Isan [northeastern Thailand] lizard") was one of the first true sauropod dinosaurs, with all four legs always on the ground. It lived 210 million years ago in Thailand. The type species is Isanosaurus attavipachi. It is known from a partial skeleton, including a neck vertebra, a back vertebra and part of another, six tail vertebra, two chevrons, fragmentary ribs, the right sternal plate, the right shoulder blade, and the left thigh bone. These remains came from a skeleton that had unfortunately been mostly eroded away. The bones belonged to an individual perhaps 6.5 metres (21 ft) long, that may not have been fully grown. They were found in dark red sandstone of the Nam Phong Formation near the village of Ban Non Thaworn, in Chaiyaphum Province.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| 05:04 pm [industrialterro] [Link] |
Herrerasaurus Герреразавр также Эрреразавр (Herrerasaurus) — один из самых примитивных динозавров подотряда тероподов. Иногда некоторые авторы считают его представителем динозавроморф — группы, включающей предков динозавров. Описан в 1963 году из позднетриасовых (карнийский ярус, 228—225 млн.л.н.) отложений Ишигуаласто в Аргентине. Назван в честь Викторио Эрреры. Самый изученный из примитивных динозавров, известен по нескольким скелетам. Современное описание на основании новых находок дал П. Серено в 1990-х годах. Внешне герреразавр напоминал более поздних хищных динозавров. Животное бипедальное, как и поздние тероподы. Передние конечности довольно длинные, четырехпалые, третий палец длиннее второго, функционально кисть была трехпалой. На задней конечности сохраняется пятая метатарзалия. По ряду признаков напоминал более поздних теропод и даже птиц. Однако имел и некоторые признаки прозауропод. Например, седалищная кость герреразавра схожа с той же костью платеозавра. Длина герреразавра достигала 3 метров (возможно, больше). Охотился на более мелких позвоночных, конкурировал с равизухидами, известными из тех же отложений. Близкородственный, но более мелкий ставрикозавр (Staurikosaurus pricei) найден в одновозрастных отложениях Бразилии. К герреразавридам может относиться также хиндезавр (Chindesaurus bryansmalli) из карнийского — норийского Техаса. Герреразавриды (Herrerasauridae), иногда эррерозавриды — семейство наиболее примитивных динозавров подотряда теропод. Герерразавриды появились на заре эпохи динозавров в позднем триасе. Это были небольшие (не больше 4 метров длиной) плотоядные ящеры, передвигающиеся на задних конечностях. Характерными особенностями строения скелета герерразавридов являются лишь частично открытая вертлужная впадина, только два крестцовых позвонка (самое маленькое количество среди диназавров). Строение лобковой кости сильно напоминает аналогичное строение у группы тетануровов (тероподов, наиболее близких к современным птицам). Шарнирно сочленённая нижняя челюсть герерразавридов похожа на челюсти всех тероподов. Довольно длинные передние конечности не имели специализированных изменений: было пять полноценных кистевых костей (в то время, как настоящие тероподы имели три длинных пальца с искривлёнными когтями), третий палец длиннее второго. Позвонки герерразавридов не имели полостей для воздушных мешков (в отличие от более поздних тероподов). Внешне герерразавриды были довольно похожими на более поздних тероподов юрского и мелового периодов, однако они не являлись их предками и полностью вымерли уже к началу юрского периода. В настоящее время семейство включает три монотипных рода Герреразавры (самый изученный род семейства, известный по нескольким скелетам), Санхуанзавры, Ставрикозавры и, возможно, ещё несколько родов (Chindesaurus, Caseosaurus), полноценная классификация которых вызывает затруднения в связи с малым количеством ископаемого материала (возможно, последние два таксона являются синонимами). Novas (1992) определил герреразавридов как Герреразавров, Ставрикозавров и их основных ближайших потомков. Другие возможные базальные тероподы: Alwalkeria и Teyuwasu. Положение герерразаврид на эволюционном дереве динозавров не вполне ясно. Они могут быть базальными тероподами, базальными ящеротазовыми или отдельной веткой, развившейся при разделении динозавров на ящеротазовых и птицетазовых. Ранние исследователи предполагали, что они представляют собой раннюю линию зауроподоморфов. Nesbitt в 2009 году предположил, что геттеразавры, и связанные с ними геттеразавриды, являются очень ранней веткой, отделившейся от тероподов, в то время как другие (например, Ezcurra, 2010) находят их базальной веткой клады Eusaurischia, что ближе к началу ветки ящеротазовых, чем к веткам тероподов или зауроподоморфов. Ситуация осложняется корреляциями слоёв окаменелостей, относимых к позднему триасу. С тероподами герреразаврид сближают некоторые характеристики — крупные когти, зубы, особенности костей таза. Многие палеонтологи, однако, не соглашались с такой классификацией и указывали на отсутствие у герреразаврид ключевых, по их мнению, признаков тероподов (к примеру, полостей в позвонках, предназначенных для воздушных мешков). Однако исследование окаменелостей Tawa hallea по всей видимости окончательно устанавливают прямую связь между тероподами и герреразавридами. Герреразавр имел длинный и узкий череп с тремя парами отверстий, исключая ноздри и глазницы. Эти отверстия уменьшали массу черепа, не снижая его прочность. Нижняя челюсть двигалась в переднезаднем направлении, благодаря гибкому соединению. По краям челюстей росли острые зазубренные зубы. Герреразавр обладал острыми зубами и охотился на мелких позвоночных животных. Сам он также являлся добычей более крупных ящеров, о чём свидетельствовали обнаруженные на его костях отпечатки зубов. Судя по зубам, герреразавр был плотоядным динозавром. Он, возможно, питался другими динозаврами типа пизанозавра, однако в те времена намного более привлекательной добычей были ринхозавры и растительноядные синапсиды. На самого герреразавра, возможно, охотились гигантские равизухиды типа заврозуха, так как на одном из черепов были найдены следы от укуса. На относительно коротких передних лапах имелась весьма длинная пятипалая кисть. Пальцы оканчивались когтями. Геррерозавр обладал сильными задними ногами и мощным хвостом, что указывало на способность быстро бегать Строение скелета этой рептили включает в себя признаки разных видов динозавров. Подвздошные кости удерживаются за счёт двух крестцовых позвонков, а лобковые кости направлены кзади, подобно дромеозавридам и птицам. Также, лобковые кости на конце имеют форму ботинка, что характерно для продвинутых теропод, а позвонки в центре похожи на песочные часы, как у аллозавра. Вследствие этого, геррерозавр долго не мог найти своей ниши в классификации древних рептилий, и эта тема постоянно была предметом научных споров. И лишь после обнаружения целого черепа, учёные пришли к общему мнению. С самого открытия этого динозавра возникли споры о его месте в классификации. Рейг считал герреразавра ранним примером карнозавров, но это вызывало много споров на протяжении последующих лет, и род классифицировался по разному. В 1970 г Стил классифицировал его как прозавропода, в 1972 г П. Гальтон посчитал его недиагностическим вне Saurischia. Несколько исследований помещали герреразавра и ставрикозавра посередине между завришиями и орнитишиями, а также вообще вне группы Dinosauria. С обнаружнием полного черепа стали преобладать два предположения - принадлежность к примитивным тероподам и группе в основе завриший, перед расхождением ее на теропод и завроподоморф. Если это тероподы, то тероподы, завроподоморфы и орнитишии развились еще раньше, чем появились герреразавриды, перед карнием, и все три линии независимо развили несколько динозавровых особенностей, типа более продвинутого соединения лодыжки и открытой вертлужной впадины. Это представление поддерживается свидетельством ископаемых следов, которые могут быть приписаны только тероподу, из ладиния формации los Rastros в Аргентине, которые предшествуют появлению герреразавра на 3-5 миллионов лет.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5, 6):
( Далее ) Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, герреразавриды, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| 04:50 pm [industrialterro] [Link] |
Halticosaurus Halticosaurus (meaning "nimble lizard") is a genus of theropod dinosaur from the Late Triassic period (middle Norian stage, around 208 million years ago). It was a coelophysoid found in the Middle Stubensandstein formation of what is present-day Germany. Some researchers consider Halticosaurus the same genus as Liliensternus, but Samuel Welles (1984) found differences in the femoral head and the anterior trochanter. The type species, Halticosaurus longotarsus, was based on partial jawbones and teeth, partial vertebrae, a few arm and leg bones, and fragments of an ilium, and was found mixed in with the remains of a prosauropod dinosaur, Sellosaurus gracilis. A second species assigned to this genus, "Halticosaurus orbitoangulatus", turned out to be a crocodylomorph (Rauhut and Hungerbuhler, 2000). A third species, H. liliensterni, has been reclassified as Liliensternus.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, целофизиды, ящеротазовые |
| 04:00 pm [industrialterro] [Link] |
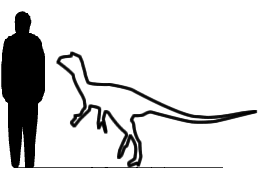
Guaibasaurus Guaibasaurus is an extinct genus of basal dinosaur known from the Triassic of Rio Grande do Sul, southern Brazil. Guaibasaurus was originally named on the basis of the holotype, MCN PV2355, a well-preserved partial postcranial skeleton and the paratype, MCN PV2356, an articulated and nearly complete left hindlimb, which were discovered in the "Sesmaria do Pinhal 2" locality near Candelária, Rio Grande do Sul, in Brazil, in the geopark of Paleorrota. Later, two additional specimens were referred to G. candelariensis: UFRGS PV0725T is an articulated and nearly complete postcranial skeleton missing one forelimb, both feet and the neck, and MCN PV 10112 is still being unprepared block containing articulated parts and some isolated elements of which, including a partial hand. The referred materials were collected from the "Linha São Luiz" locality near the town of Faxinal do Soturno, Rio Grande do Sul, also in the geopark of Paleorrota. All specimen were collected in these two localities from the lower portion of the Caturrita Formation (Rosário do Sul Group, Paraná Basin) or alternatively the uppermost Santa Maria 2 Sequence, dating to the early Norian faunal stage of the Late Triassic, about 216.5-212 million years ago. Specimen UFRGS PV0725T is articulated with hindlimbs tucked underneath its body and forelimbs flexed to the side. Although most of the neck is not preserved, the vertebrae at the base of the neck are present in UFRGS PV0725T and curve to the left, suggesting the entire neck was curved toward the left side of the body. The posture of this skeleton is similar to the resting position of birds, and is otherwise known only from advanced maniraptoran dinosaurs that are closely related to birds. Like living birds, Guaibasaurus may have rested in this position to conserve body heat. José Bonaparte and colleagues, in their 1999 description of the genus, found it to be possible basal theropod and placed it in its own family, Guaibasauridae. Bonaparte and colleagues (2007) found another early Brazilian dinosaur Saturnalia to be very similar to it, and placed the two in the Guaibasauridae which was found to be a primitive saurischian group. Bonaparte found that these forms may have been primitive sauropodomorphs, or an assemblage of forms close to the common ancestor of the sauropodomorphs and theropods. Overall, Bonaparte found that both Saturnalia and Guaibasaurus were more theropod-like than prosauropod-like. However, all more recent cladistic analyses found the members of Guaibasauridae to be very basal sauropodomorphs, except Guaibasaurus itself which was found to be a basal theropod or alternatively a basal sauropodomorph. Guaibasauridae — семейство примитивных ящеротазовых динозавров, известных по ископаемым останкам из позднего триасового периода на территории современных Бразилии и Аргентины. Точный состав и классификация Guaibasauridae остается неопределенной. Семейство первоначально было введено Хосе Бонапартом и его коллегами в 1999 году, тогда содержало один род и один вид — Guaibasaurus candelariensis, и было первоначально причислено к группе тероподов. После второго найденого образца Guaibasaurus, этот динозавр был описан более подробно в 2007 году, что дало возможность сравнить его с другими малоизученными представителями ранних ящеротазовых динозавров, которых часто трудно классифицировать, поскольку они сочетают в себе характеристики двух основных групп ящеротазовых — тероподов и зауроподоморфов. Бонапарт и его коллеги, в свете новой информации, полученной от второго образца, пришли в выводу, что род Saturnalia (который анатомически очень похожи на Guaibasaurus) может быть также отнесен к Guaibasauridae. При дальнейших исследованиях Бонапарт с коллегами обнаружили, что Guaibasauridae обладают большим количеством черт, присущих тероподам, нежели чем черт, присущих ранним зауроподоморфам (или «прозауроподам»). Таким образом, согласно результатам исследований Бонапарта, Guaibasauridae, скорее всего, являются очень ранней базальной группой, находящейся на вершине филогенетической схемы, ведущей к завроподоморфам или группе общих предков зауроподоморфов и тероподов. Кроме того, авторы интерпретируют эти данные, как доказательство того, что общий предок обеих линий ящеротазовых динозавров по внешнему виду был более близок к тераподам, чем к прозауроподам. По классификации M. D. Ezcurra (2010) семейство включает в себя подсемейство Saturnaliinae, содержащее 2 рода — Chromogisaurus и Saturnalia.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, тероподы, ящеротазовые |
| 03:48 pm [industrialterro] [Link] |
Gojirasaurus Gojirasaurus (meaning "Godzilla Lizard") is a dubious genus of dinosaur named after the giant monster movie character Gojira (the Japanese name for Godzilla). It was discovered in the Bull Canyon Formation (Dockum Group of Norian age, near Revuelto Creek, Quay County, New Mexico). The type specimen is a partial skeleton, estimated to be about 5.5 m (18 ft) long, which can be extrapolated to a weight of approximately 150–200 kg (330–440 lb). It lived during the middle Norian age of the late Triassic Period, about 210 million years ago. Gojirasaurus was a member of the Coelophysoidea, which were small, early theropods. Gojirasaurus is one of the largest meat-eating dinosaurs known from the Triassic Period, and was enormous for its time. The original remains attributed to this dinosaur included a serrated tooth, four ribs, and four vertebrae, hip bones and a tibia (lower leg bone). However, later study showed that the vertebrae actually belonged to rauisuchian Shuvosaurus. Today, only the tibia and hip bone can be attributed to a coelophysoid dinosaur. The leg and hip bones are indistinguishable from the contemporary Coelophysis, making its status as a valid genus dubious. In early, unpublished studies which included this specimen, the dinosaur was referred to by the name "Revueltoraptor lucasi", a nomen nudum.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, целофизиды, ящеротазовые |
| 03:29 pm [industrialterro] [Link] |
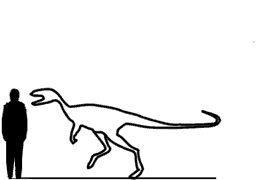
Eoraptor Эора́птор (Eoraptor lunensis) — один из наиболее ранних представителей группы динозавров, обитавший в позднем триасе в северо-западной области Аргентины 228—225 миллионов лет назад. Окаменелости этого динозавра впервые были обнаружены в 1991 году палеонтологом Чикагского университета Полом Серено, в Аргентине, в месте Валье де ла Луна (исп. Valle de La Luna, Долина Луны). Позже было обнаружено ещё несколько хорошо сохранившихся скелетов. Название типового вида означает «лунный рассветный грабитель» (др.-греч. eos/εως означает «утро» или «заря», англ. raptor — «грабитель», лат. lunensis — «лунный», «с Луны»). Последняя часть названия отражает место первого нахождения останков этого динозавра. Тело располагалось вертикально на задних ногах, а его передние лапы были в два раза короче задних и имели по пять «пальцев». Имел размеры приблизительно 1 метр в длину и массу приблизительно 10 килограммов. Строение скелета позволяет причислить эораптора к отряду ящеротазовых динозавров, однако более точное таксономическое положение рода неясно. Возможно, эораптор относится к тероподам. В то же время определённые детали его строения (к примеру, увеличенные ноздри) характерны для растительноядных зауроподоморфов. Пожалуй, самым тривиальным из всех известных нам динозавров является эораптор. Его нашли в 1993 году на месторождении позднего триасового периода, а именно в пустошах бассейна Ишигуаласто, что в северозападной Аргентине. Его отыскали Пол Серено и Фернандо Нова вместе с коллегами на том же месторождении, где ранее отыскали геррерозавра, который является ещё одним достаточно примитивным динозавром. Эораптор был найден почти случайным образом. Один учёный из команды собирался отбраковать одну из горных пород, и вдруг заметил, что внутри нее находятся зубы. Он рассмотрел её поближе и, оказалось, что в этой породе находится цельный череп. В результате этой находки ареал стали штудировать дальше и раскопали цельный скелет эораптора – динозавра который до этого не был известен. Его возраст был почти 230 000 000 лет. Судя по этому скелету, можно с полной ответственностью заявить, что эораптор являлся двуногим хищным животным, очень примитивным родственником тероподов. Он перемещался, в основном, на задних конечностях, но временами мог перемещаться и на всех четырех. Хотя на «руке» у него и было пять пальцев, один из них был очень маленьким. Как и у остальных тероподов, у эораптора были тонкостенные и полые кости конечностей, на стопе было три длинных пальца. Вместе с тем, отличие от остальных тероподов, у эораптора первый палец ноги мог являться опорным при перемещении. Зазубренные зубы эораптора свидетельствовали о том, что он являлся плотоядным животным, а в своих цепких лапах передних конечностей мог удерживать жертву, которая по размерам не уступала ему самому.
Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, ящеротазовые |
| March 17th, 2012 | |
| 06:41 pm [industrialterro] [Link] |
Eodromaeus Эодромеус (Eodromaeus murphi) — ископаемый плотоядный динозавр из группы тероподов, живший около 230 миллионов лет назад во время позднего триаса, в районе нынешней Аргентины. Находки были сделаны в предгорьях Анд в 1996 году. Классифицируя древнего ящера, учёные поместили его у «корней» эволюционного дерева тероподных динозавров. Общая длина тела животного, по данным учёных, — 1,2 м, а масса взрослой особи доходила до 5 кг. Изучение E. murphi заставило палеонтологов пересмотреть свои взгляды на классификацию эораптора, предлогая поместить его в группу зауроподоморфов, ссылаясь на определённые детали его строения (к примеру, увеличенные ноздри) характерные для данной группы. Американские и аргентинские палеонтологи описали новый вид хищных динозавров – один из древнейших на сегодняшний день. Этот динозавр, получивший название Eodromaeus murphi, может считаться одним из самых древних тероподных динозавров, на эволюционном древе теропод он располагается у самого основания ствола. Он жил в начале позднего триаса, в карнийском веке – 230 миллионов лет назад. Поэтому-то его и назвали Eodromaeus, что можно перевести как "рассветный бегун" - он бегал по земле на юго-западе древнего континента Пангеи на заре эры динозавров. Коллектив палеонтологов под руководством Риккардо Мартинеса (Ricardo N. Martinez) из Университета Сан-Хуана (Universidad Nacional de San Juan) и Пола Серено (Paul C. Sereno) из Университета Чикаго (University of Chicago), опубликовал статью, посвященную описанию Eodromaeus murphi, в журнале Science.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| 06:09 pm [industrialterro] [Link] |
Eocursor Эокурсор (Eocursor; букв. «ранний бегун») — род вымерших птицетазовых динозавров, живших в позднем триассовом периоде (около 216,5 - 203,6 миллионов лет назад), на территории нынешней Южной Африки. Впервые описан палеонтологом Ричардом Батлером (Richard Butler) и его коллегами в 2007 году. Представлен одним видом - Eocursor parvus. Название рода Eocursor образовано от греческого слова «eos» – «рассвет» или «начало» и латинского «cursor» – «бегун». Видовое название образовано от латинкого «parvus» – «маленький». Окамнелые останки Eocursor были найдены в 1993 году на территории фермы Damplaats (55), близ города Ladybrand, ЮАР, в геологической формации Lower Elliot относящейся к норианскому ярусу. Голотип SAM-PK-K8025 состоит из частичного скелета. Реконструированый внешний вид, а также описание динозавра было представлено только в 2007 году палеонтологами R. Butler, R. Smith и D. Norman. Принадлежит к плохо изученным на сегодняшний день ранним птицетазовым динозаврам (Ornithischia). По внешнему виду сильно отличается от общепринятого облика рептилий данного отряда. Искомпаемые останки включают в себя кости черепа, конечностей, таза и позвоночника, и являются наиболее полными известными останками скелета птицетазового динозавра триасового периода на территории Южной Африки. В длину Eocursor parvus достигал 1 метра, а высота не превышала 30 см. Листообразные зубы ящера предназначались для пережевывания исключительно растительной пищи. Верхние конечности, обладавшие большими, несоизмеримыми с телом, размерами, были очень удобными для хватания. А строение костей ящера и их небольшой вес свидетельствует о том, что животное могло быстро и проворно перемещаться. Птицетазовые (Ornithischia) — один из двух отрядов динозавров. Несмотря на своё название, не они, а ящеротазовые динозавры по одной из версий были предками птиц. Были травоядными (за исключением ранних всеядных видов), меньших размеров, чем ящеротазовые зауроподы. Кроме самых примитивных, имеют приспособления к питанию растительной пищей: сложный кишечник и щёки. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Таз имел четырёхлучевое строение. Лобковая кость имела вырост, направленный назад вдоль седалищной кости (у ящеротазовых кости были практически перпендикулярны). Лобковая кость часто образует новый вырост, направленный вперёд параллельно позвоночнику. Подобное строение таза развивалась у динозавров независимо трижды: у птицетазовых, у надсемейства Therizinosauroidea и семейства Dromaeosauridae. Последнее и является наиболее вероятным предком птиц. У некоторых видов (Hypsilophodon, Talenkauen и Thescelosaurus) имеются межреберные костные пластины в передней части ребер. Некоторые исследователи предполагают, что эти пластины выполняли биомеханические функции, помогали в дыхании либо были предназначены для обеспечения повышенной жесткости передней части грудной клетки, в то время как другие полагают, что они служили для оборонительных функций. Анализ имеющихся у ученых останков птицетазовых динозавров, позволил им определить, что ящеры данного отряда впервые появились в позднем триасе, примерно 220 млн лет назад и вначале были крайне малочисленны. Этот отряд динозавров начал бурно эволюционировать только в последующий юрский период (208–144 млн лет назад), все больше распространяясь по планете и занимая экологические ниши после исчезновения иных травоядных существ. Ранее птицетазовые делились на 4 или 6 подотрядов, включая тиреофоров (иначе (стегозавров), рогатых динозавров (цератопсов), анкилозавров (панцирных динозавров), цераподов (известнейшие представителей которых — игуанодоны) и панцироголовых пахицелозавров. В настоящее время они подразделяются на два подотряда-клады: тиреофоров и цераподов. Первые включают инфраотряды стегозавров и анкилозавров, вторые — рогатых динозавров и пахицефалозавров (со своей стороны нередко объединяемых в кладу или подотряд маргиноцефалов), и орнитоподов (включая игуанодонов, утконосых гадрозавров и др.)
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, птицетазовые |
| 05:37 pm [industrialterro] [Link] |
Efraasia Efraasia — род вымерших ящеротазовых динозавров из группы зауроподоморфов (Sauropodomorpha), живших в позднем триасовом периоде (около 210 миллионов лет назад), на территории нынешней Европы. Окаменелости динозавра были найдены в Германии. Впервые описан палеонтологом F. von Huene d 1907–1908 под названием Teratosaurus minor. Представлен одним видом - E. minor. Efraasia was once thought to be a relatively small dinosaur, about 2 to 3 metres (6.6 to 9.8 ft) long, but this was because the most complete known fossils are from juvenile animals. Yates in 2003 has estimated the adult length at 6.5 metres (21 ft); the largest specimen is SMNS 12843 with a femur length of 627 millimetres (24.7 in). Efraasia was lightly built for its size, with gracile hands and feet. Like many "primitive" sauropodomorphs, Efraasia might have been partially bipedal and partly quadrupedal. It had long fingers and mobile thumbs, with which it would have been able to grasp food, but the shape of its wrists might have allowed it to walk easily on all fours. Some researchers however, contend that the lower arm did not allow pronation, a rotation of the radius around the ulna, so that the hand could not be directed downward, making the animal an obligate biped. The skull is small, pointed and triangular. There are four teeth in the premaxilla. The neck is only moderately elongated but thin. The neural spines of the tail are low. The second finger is longer than the third finger. The first toe is not strongly reduced. Von Huene identified a cluster of stomach stones (gastroliths) in association to specimen SMNS 12667. Yates identifies two unique derived traits (autapomorphies): the presence of a raised crescent-like ridge on the upper part of the inner side of the pubis shaft; and the presence of a vaulted bony web between two lower extensions of a braincase bone, the processus basipterygoidei, with a raised central bony platform on top of the vault. Von Huene continued interpreting these forms as predatory dinosaurs, in 1932 assigning them to a separate family Palaeosauridae as part of the Carnosauria. Only in 1965 Charig established they were plant-eating sauropodomorphs. In 1973 Galton assigned Efraasia to the Anchisauridae, but he used this name as a paraphyletic group encompassing all "prosauropods" that were not melanorosaurids. Modern phylogenetic analysis has indicated that Efraasia is a basal sauropodomorph, somewhat more derived than Thecodontosaurus, but less than either the Prosauropoda (including Plateosaurus) or the Sauropoda. The genus is sometimes recovered as the sister taxon to the last common ancestor of both larger groups.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, ящеротазовые |
| 05:10 pm [industrialterro] [Link] |
Daemonosaurus Демонозавр (Daemonosaurus) — род ящеротазовых динозавров из группы тероподов, живших в позднем триасовом периоде 205 млн лет назад на территории нынешней Мексики. Окаменелости теропода были найдены в геологической формации Chinle Formation в карьере Ghost Ranch штата Нью-Мехико. Впервые описан группой палеонтологов во главе с Hans-Dieter Sues в 2011 году. Представлен одним видом - Daemonosaurus chauliodus. По данным кладистического анализа останков, Daemonosaurus находится в базальных тероподах, находясь в генеалогическом древе чуть ниже эораптора и выше Tawa и вне группы Neotheropoda (группы, которая включает в себя более продвинутых триасовых тероподов, таких как Coelophysis и их потомков). С таким базальным положением, он открывает новую линию развития, которая простирается от ранних форм динозавров среднего триаса, таких как Eoraptor и Herrerasaurus из Южной Америки. Филогенетический анализа показал его тесную связь с Tawa hallae. Хотя эти двое тероподов тесно связаны друг с другом, Tawa был найден в каменоломне, которая немного старше, чем Ghost Ranch. Нового, ранее неизвестного науке динозавра обнаружили американские палеонтологи. Череп с острыми зубами, несколько шейных позвонков и обломки ребер нашли в так называемом Карьере целофизиса (Coelophysis Quarry) в районе Ранчо Призрака (Ghost Ranch) в штате Нью-Мексико. Этот район со зловещим названием хорошо известен палеонтологам, там неоднократно находили скелеты новых видов динозавров, живших в триасовом периоде. Сам карьер назван в честь триасового динозавра целофизиса (Coelophysis), кости которого там встречаются довольно часто. Находка этого динозавра интересна не только тем, что обнаружен новый вид триасовых теропод, которых известно не так много, но и тем, что он оказывается своего рода "промежуточным звеном" между уже известными видами. В 2009 году "Аммонит.ру" рассказывал об обнаружении в районе того же самого Ранчо Призрака другого триасового теропода - Tawa hallae. Этот динозавр был очень важным открытием – он позволил связать древних хищных геррерозавров (Herrerasaurus), еще не очень похожих на юрских теропод, с более молодыми целофизисами (Coelophysis), уже вполне тероподными динозаврами. Новый Daemonosaurus на эволюционном древе ранних теропод располагается между геррерозаврами и Tawa hallae, в его черепе и позвонках палеонтологи рассмотрели как базальные (свойственные более древним видам), так и продвинутые признаки. Правда, предком Tawa демонозавр не был, его кости найдены в несколько более молодых отложениях, чем кости Tawa. Следовательно, демонозавры были «боковой ветвью», отошедшей от общего ствола ранних теропод несколько раньше, чем появились Tawa hallae. Несмотря на относительную примитивность строения, они успешно конкурировали со своими более молодыми родственниками и, судя по всему, жили одновременно с целофизисами в самом конце триаса.
Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, ящеротазовые |
| March 16th, 2012 | |
| 07:35 pm [industrialterro] [Link] |
Coelophysis Целофизис, также целофиз (Coelophysis) — род небольших хищных динозавров подотряда тероподов, живщих в конце триасового периода (около 215 миллионов лет назад) на территории нынешней Северной Америки. Типовой вид — Coelophysis bauri. Размеры целофиза достигали от двух до трёх метров в длину, полтора метра в высоту. Весил 15—30 килограммов. Своё название целофизис получил от Эдварда Копа в 1889 году, что означает «пустые формы» со ссылкой на его полые кости. Coelophysis обладал очень изящным телом, что вероятно свидетельствует о его способности хорошо бегать. Несмотря на то, что он является ранним динозавром, строение его тела уже существенно отличалось от других ранних динозавров, таких как Herrerasaurus и Eoraptor. Туловище Coelophysis соответствует тероподам, но грудной пояс показывает некоторые интересные особенности: у Coelophysis bauri была так называемая вилочка (furcula), самый ранний известный пример у динозавров. Coelophysis также обладал четыремя пальцами на передних конечностях. Целофизисы были опасными хищниками. Они охотились на ящеров крупнее них, и несомненно питались падалью. До начала 2000-х годов высказывалось предположение, что целофиз был каннибалом, основанное на том, что в Ghost Ranch был найден скелет детеныша целофизиса в желудке взрослого динозавра. Тем не менее, Боб Гей в 2002 году высказал мнение, что эти образцы были неверно истолкованы (несколько экземпляров «детенышей целофизиса» на самом деле являются останками небольших рептилий, таких как Hesperosuchus), и что больше нет каких-либо доказательств в поддержку теории о каннибализме у целофиза. Результаты исследования Боба Гея были подтверждены в 2006 году последующими исследованиями Несбитта и его коллег. Новые находки содержимого желудков целофизов могли бы внести ясность в данный вопрос. Происхождение карнозавров почти неизвестно, зато ученым удалось узнать, что эволюционировали они быстро. К середине юры большинство хищных динозавров составляли именно карнозавры. Одними из самых первых динозавров были мелкие, изящные, тонконогие целофизиды, появившиеся около 210-200 млн л. н. Известнейший вид целофизидов - целофизис. Жил он в самом начале эры динозавров в западной части Северной Америки. В Нью-Мексико найдено огромное количество исклпаемых останков этих проворных ящеров. Можно с уверенностью сказать, что целофизис был плотоядным. Вероятно, он питался небольшими животными: ящерицеобразными рептилиями, амфибиями, насекомыми, рыбой. У него было довольно много острых как бритва зубов, но они были недостаточно велики, чтобы разрывать крупную добычу. Целофизисы были одними из первых динозавров на Земле. И одними из первых они были досконально изучены. Несомненно, эти ящеры были опасными хищниками, пусть и не такими большими, как их позднейшие родичи, хотя на пустынных пространствах триаса, особенно во время засух им, пожалуй, равных не было. При нападении на жертву целофизисы пускали в ход свои передние конечности. Вполне вероятно, что такие ящеры, как целофизис, были предками первых карнозавров, появившихся на Земле на миллионы лет позже. При эволюционном переходе от целофизиса к карнозавру изменения в общей форме тела и его положении при ходьбе должны были быть минимальными. Главные же различия заключаются в том, что целофизиды имели более хрупкое телосложение, нежели чем карнозавры: с тонкими конечностями (причем передние конечности почти не уступали в размерах задним), тонкой длинной шеей и довольно маленькой узкой головой. Отдельные кости скелета целофизидов по весу очень легкие, с тонкими стенками, да еще и с дополнительными, снижающими общий вес, "окнами" в полости. А вот у большинства ранних карнозавров кости крупнее, мощнее, плотнее, тяжелее. Тем не менее вышесказанное является лишь предположением. Ведь целофизис жил около 210 млн л. н., а возраст большинства самых ранних ископаемых остатков карнозавров составляет примерно 170-160 млн лет. Кроме того, ископаемых, свидетельствующих о существовании переходных форм от целурозавра к карнозавру в промежутке времени между двумя этими периодами, пока не найдено. Целофизи́ды (Coelophysidae) — семейство примитивных хищных динозавров подотряда тероподов. Большинство видов данного семейства были относительно небольшими по размеру. Процветали в позднем триасе и раннем юрском периоде на территории Северной Америки, Африки и Азии. В кладистике, целофизиды были первоначально определены Полом Серено в 1998 году как группа, произошедшая от общего предка целофиза (Coelophysis bauri) и прокомпсогната (Procompsognathus triassicus), и включающая в себя всех потомков общего предка. Целофизиды - часть клады Coelophysoidea. Старый термин «Podokesauridae», введеный 14 годами ранее до целофизидов (и имевший больший приоритет, чем нынешний термин), в настоящее время обычно игнорируется, так как его типовый экземпляр был разрушен в огне и больше не может быть использован для сравнения с новыми находками. Теропо́ды (Theropoda, от греч. θηρίο, «зверь», и πόδι, «нога», — звероногие), или хищные динозавры — один из подотрядов ящеротазовых динозавров. Все тероподы перемещались на двух ногах, большинство — хищники, реже всеядные или растительноядные (теризинозавры, орнитомимиды). Большинство ученых с недавних пор стали относить к тероподам и современных птиц, таким образом птицы из потомков динозавров стали относиться к одному из подотрядов. Кисть обычно трёхпалая, стопа — обычно четырёхпалая с опорой на 2—4-й пальцы. Крупнейшим среди тероподов был спинозавр (более 17 метров). Тероподы составляли около 37 % всех известных родов динозавров, среди последних находок Nemegtomaia (2004), Гуанлонг (2006), Limusaurus (2009) и др. Эволюционное происхождение перьев можно проследить начиная с Carnotaurus покрытого многочисленными пупырышками, похожими на фолликулы. Следующий этап виден у Sinosauropteryx и Dilong покрытых волокнистым пухом. И наконец у Caudipteryx, Protarchaeopteryx, Sinornithosaurus, и Microraptorа можно наблюдать настоящие перья. Считается, что примерно 230 миллионов лет назад, когда все континенты были объединены в суперконтинент Пангею, древние ящеры стали заселять разные регионы планеты. Тероподы возникли на территории Гондваны (региона, который позже стал Южной Америкой) и позже расселились по суперконтиненту. Известно, что к концу триасового периода (около 228—216.5 млн лет), три основных линии динозавров, а именно тероподы, завроподоморфы и птицетазовые были уже хорошо дифференцированы. Находки ранних тероподов, таких как Herrerasaurus, Staurikosaurus, Eoraptor, Eodromaeus и Sanjuansaurus были найдены в Южной Америке на территории нынешних государств Аргентины и Бразилии. Эти примитивные тероподы имели пять пальцев на обеих руках и ногах (хотя и с значительно уменьшенным пятым пальцем), в то время как более позднии тероподы из группы — Neotheropoda, имели только три пальца на своих руках (у некоторых третий палец был рудиментарным), и три функциональных пальца на ногах. До недавнего времени, стадии более позднего триаса (216—199 миллионов лет назад) были представлены в основном только одной группой Coelophysoidea, принадлежащей к неотераподам (куда включены Coelophysis, Segisaurus, Lilensternus и возможно Zupaysaurus). Эволюционный разрыв между ранними тероподами и неотераподами начал заполняться начиная с 2009 года, с описанием новых видов — Tawa hallae, Daemonosaurus. В результате анализа палеонтологических данных было установленно, что тероподы поэтапно мигрировали из Южной в Северную Америку; перемещались с места на место, как и другие группы животных (к примеру, крокодиломорфы). Эти находки доказывают, что уже примерно 215 млн лет назад терподы заселили территорию современного штата Нью-Мехико - например динозавр Tawa.
Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9):
( Далее ) Размеры тела в сравнении с человеком (закрашен фиолетовым цветом):
Ископаемые останки (1, 2, 3, 4, 5, 6, 7, 8):
( Далее ) Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, целофизиды, ящеротазовые |
| 07:10 pm [industrialterro] [Link] |
Chromogisaurus Хромогизавр (Chromogisaurus) — ископаемый род примитивных ящеротазовых динозавров из семейства Guaibasauridae, живших в конце триасового периода (около 228.7—216.5 миллионов лет назад) на территории нынешней Южной Америки. Представлен одним видом — Chromogisaurus novasi. Окаменелости зауроподоморфа (голотип PVSJ 846) были найдены в геологической формации Ischigualasto Formation относящейся к Carnien (провинция Сан Хуан в Аргентине). Впервые описан палеонтологом Мартином Д. Эскуррой (Martín D. Ezcurra) в 2010 году. Известен по частичному посткраниальному скелету. Он включает в себя правую локтевую кость, весь таз, кости бедра, левой ноги, правой голени, фаланги, среднего и переднего хвостового позвонка. Название рода происходит от др.-греч. «chroma» — «цвет», и «gè» — «страна», ссылаясь на Лос-Колорадос, красочные скальные образования в Valle Pintado. Видовое название дано в честь аргентинского палеонтолога Фернандо Эмилио Новаса (Fernando Emilio Novas). Chromogisaurus является одним из старейших известных динозавров. В длину достигал около 2 метров. По словам исследователей форма относительно длинной локтевой кости указывает на то, что эти динозавры хотя бы иногда передвигались на четырёх конечностях. Задние конечности значительно длиннее передних. В результате кладистического анализа Мартин Эскурра отнес Chromogisaurus к базальным зауроподоморфам в семейство Guaibasauridae, вместе с Guaibasaurus, Agnosphitys, Panphagia и Saturnalia. В Guaibasauridae образует небольшое подсемейство Saturnaliinae вместе с сестринским таксоном Saturnalia.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, ящеротазовые |
| 06:41 pm [industrialterro] [Link] |
Chindesaurus Chindesaurus bryansmalli — ископаемый вид ящеротазовых динозавров, живших в позднем триасе около 216 млн лет назад на территории США. Находки были сделаны в штате Аризона. В настоящее время точное таксономическое положение динозавра не определено, из-за фрагментарного характера его останков. Предполагается, что он относится к кладе Herrerasauridae. Если это действительно так, то он является самым поздним представителем данной группы. Chindesaurus is known from five incomplete specimens (six if Caseosaurus is included). The type specimen is the most complete, and contains several rib and vertebrae fragments, two complete vertebrae from the hips and one from the tail, fragments of hip bones, and partial leg bones. A single tooth was also found with the specimen. The remaining specimens are even more incomplete, consisting of isolated hip bones and upper leg bones (femora), and more vertebrae. The type and paratype specimens probably came from individuals around 2.4 metres (7.9 ft) in length. Chindesaurus has been difficult to classify, and has been recovered in several different positions at the base of the saurischian family tree. When it was first discovered in 1985, the animal which would eventually be named Chindesaurus was thought to be a prosauropod. When it was finally described and named a decade after its discovery by Long and Murry, they regarded it as a herrerasaurid, an opinion that has been followed by most paleontologists since. Phylogenetic analyses published through 2007 continued to find it to fall among the herrerasaurids, though a few studies cast doubt on this, including one in 2007 by Irmis, Nesbitt and colleagues which found Chindesaurus to be a probable basal saurischian dinosaur, and noted that it shares a wide range of characteristics with several lineages of basal saurischians, making any classification problematic. One specimen originally assigned to Chindesaurus, from the Tecovas Formation of Texas, was later placed in its own genus and species, Caseosaurus crosbyensis. Subsequent research has shown that this separation was probably in error, and that the two forms represent the same species. Nesbitt, Irmis and Parker agreed in a 2007 paper that there is little reason to separate Caseosaurus from Chindesaurus, and the two even share some unique characteristics not found in similar species. Nesbitt and colleagues suggested that any differences between the two were probably related to differences in size. However, since both species are so fragmentary, they decided not to formally make them synonyms.
Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4):
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, ящеротазовые |
| March 12th, 2012 | |
| 05:36 pm [industrialterro] [Link] |
Antetonitrus Antetonitrus (pronounced ant-EE-tohn-IET-rus; meaning "before the thunder") is the oldest known genus of sauropod dinosaur, living during the Late Triassic Period of southern Africa. It was a quadrupedal herbivore, like many of its later relatives, although it was far smaller than some of them. Antetonitrus was the largest animal in its environment, reaching up to 33 feet (10 m) long and weighing up to two tons, but still shows some primitive adaptations to use the forelimbs for grasping, instead of purely for weight support. The fossils now known as Antetonitrus were actually discovered by Kitching in 1981 in the Free State of South Africa, and were stored in the Bernard Price Institute where they were labeled as Euskelosaurus. Yates recognized them as a separate taxon and published a description several years later. The holotype, or original specimen, consists of several vertebrae and numerous bones from both forelimb and hind limb, all presumed to be from one individual. Five more limb bones from another smaller individual were also referred to the genus. Antetonitrus shows several features which appear to be approaching those of sauropods, but still retains some primitive features. Unlike most of its smaller and more lightly built ancestors, Antetonitrus was primarily quadrupedal. Like sauropods, its forelimbs were much longer relative to its hind legs than earlier animals, and the wrist bones were broader and thicker to support more weight. However, the first digit of the hand, also called the "thumb" or pollex, was still twisted and flexible, capable of grasping against the hand. In more derived sauropods, the wrist bones are large and thick, arranged in such a way as to lock the hand into a permanently pronated position for full-time weight support, and the hand is incapable of grasping. A cladistic analysis recognizes Antetonitrus as a basal sauropod, occupying a position between more derived animals such as Isanosaurus or Vulcanodon, and more basal sauropods like Melanorosaurus. The back vertebrae are extremely similar to Lessemsaurus from South America, while the limb bones are similar to Blikanasaurus, another stocky early sauropod from South Africa. However, these animals were not included in a cladistic analysis with Antetonitrus because they are poorly known (Yates & Kitching, 2003). While Antetonitrus is not the earliest sauropod from a phylogenetic standpoint, it currently the oldest known sauropod chronologically, or rather tied for that distinction with other early sauropods from the same formation, like Melanorosaurus and Blikanasaurus. Fossils of these animals were recovered from the Lower Elliot Formation, which dates to the Norian stage of the Late Triassic, or approximately 221 to 210 million years ago. Before Antetonitrus and the other Lower Elliot animals were recognized as sauropods, the oldest known sauropod had been Isanosaurus from the Rhaetian stage, a slightly younger segment of the Triassic, of Thailand (Buffetaut et al., 2000). Early sauropods and their prosauropod relatives were found around the world as all of the continents were at the time united into the single supercontinent, Pangaea, which made dispersal across the entire terrestrial world possible.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, ящеротазовые |
| 05:21 pm [industrialterro] [Link] |
Alwalkeria Alwalkeria ( This dinosaur was originally named Walkeria maleriensis by Sankar Chatterjee in 1987, in honor of famous British paleontologist Alick Walker and the Maleri Formation, in which its fossils were found. However, the original generic name was found to be preoccupied by a bryozoan. A new name was created in 1994 by Chatterjee and Ben Creisler. Remains of Alwalkeria have been recovered from the Maleri Formation of Andhra Pradesh, India. This is a Late Triassic geologic formation. Indeterminate prosauropod material has also been found in the Maleri, but Alwalkeria is the only named dinosaur species. The one known specimen is partial and consists of parts of the front ends of the upper and lower jaws, 28 incomplete vertebrae from all parts of the spinal column, most of a femur (thigh bone), and an astragalus (ankle bone). The partial skull is about 4 centimeters long (1.5 in). Although material of Alwalkeria is limited, the spacing and shape of the teeth strongly resemble those of Eoraptor. As in Eoraptor, a gap separates the teeth of the premaxillary and the maxillary bones of the upper jaw. Other similarities in the skull of the two animals also link them on morphologic grounds (Langer 2004). Alwalkeria has not been included in a cladistic analysis, but its similarities to Eoraptor suggest it may have held a similar position in the dinosaur family tree. However, the position of Eoraptor is disputed. One recent analysis finds it within the order Saurischia, but basal to the Theropoda-Sauropodomorpha split (Langer 2004). Paul Sereno finds instead that Eoraptor is a basal theropod (Sereno 1999). Others place Eoraptor outside of Dinosauria completely (Fraser et al. 2002). This dinosaur also has a heterodont dentition in the upper jaw, meaning that the teeth are differently shaped depending on their position within the jaw. Similarly to Eoraptor and basal sauropodomorphs, the front teeth are slender and straight, while the teeth in the sides of the jaw are curved backwards like those of predatory theropods, although none are serrated. This arrangement of teeth is neither clearly herbviorous nor clearly carnivorous, which suggests that this dinosaur was an omnivore with a varied diet, including insects, small vertebrates, and plant material. Several features make Alwalkeria unique among basal dinosaurs. Besides its unserrated teeth, and the mandibular symphysis is proportionally wider than almost any other known dinosaur. Also, there is a very large articulation between the fibula and the ankle.
Tags: Вымершие рептилии, Триас, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, ящеротазовые |







































































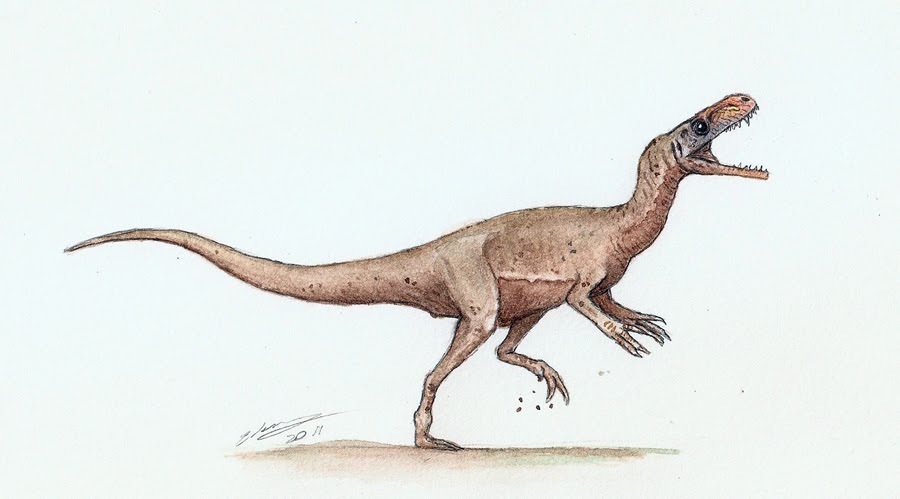






































{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.png){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}