[Recent Entries][Archive][Friends][User Info]
Below are 20 entries, after skipping 20 most recent ones in the "Сообщество, посвящённое ра" journal:[<< Previous 20 entries -- Next 20 entries >>]
| February 18th, 2015 | |
|---|---|
| 06:23 pm [industrialterro] [Link] |
Agathaumas Агатаум (от Agataumas — «огромное изумление») — сомнительный род динозавров семейства Цератопсиды, живших 70—65 млн лет назад (середина кампанского — конец маастрихского яруса позднего мелового периода) на территории нынешней Северной Америки. Достигал в длину 9 метров, высоты 3 метра и веса 5 тонн. Окаменелости агатаума были найдены в округе Суитуотер (Sweetwater County) штата Вайоминг. Впервые описан палеонтологом Коупом в 1874 году. Представлен одним видом — Agathaumas sylvestris. Останки агатаума обнаружил американский учёный Ф. Меек в 1873 году при раскопках в штате Вайоминг (США). Он сообщил об этом известному палеонтологу Эдварду Коупу, который немедленно прибыл на место и занялся изучением находки. Название своё — «огромное изумление» — агатаум получил потому, что учёные были сильно потрясены размерами животного. В 1897 году художник Чарльз Найт по заказу Э. Коупа сделал ряд рисунков ящера, которые в 1925 году послужили Уиллису О’Брайну основой для создания образа агатаума в фильме «Затерянный мир». Agathaumas (/æɡəˈθɔːməs/; "great wonder") is a dubious genus of a large ceratopsid dinosaur that lived in Wyoming during the Late Cretaceous (late Maastrichtian stage, 66 million years ago). The name comes from Greek, αγαν - 'much' and θαυμα - 'wonder'. It is estimated to have been 9 metres (30 ft) long and weighed 6 tonnes (5.9 long tons; 6.6 short tons), and was the largest land animal known at the time of its discovery. It was the first ceratopsian known to science, though relatively little is known about it. The original specimen consisted only of the animal's hip bones, hip vertebrae and ribs, and because these bones vary little between ceratopsid species, it is usually considered a nomen dubium. It is provisionally considered a synonym of Triceratops, but is difficult to compare to that genus because it is only known from post-cranial remains. The fossil remains of Agathaumas were first found in 1872 in southwestern Wyoming. They were discovered by Fielding Bradford Meek and H.M. Bannister while they were looking for fossil shells in the Lance Formation (then Laramie Formation) near the Black Butte and Bitter Creek. Meek and Bannister were employed by Ferdinand Hayden's Geological Survey of the Territories, and notified paleontologist Edward Drinker Cope of the find. Cope himself searched the ridge near Black Butte and re-discovered Meek's site, finding huge bones protruding from the rocks near a coal vein. The bones were preserved in sand and clay sediments, packed with fossil sticks and leaves, indicating a heavily forested habitat. Cope later (in 1873) described the skeleton as "the wreck of one of the princes among giants." Later in 1872, Cope published a description and name for the animal, Agathaumas sylvestris, or "marvelous forest-dweller," in reference to its great size and the environment revealed in the same rocks as its bones. The name Agathaumas has been cited as an example of Cope's excitement with this discovery, which was, at the time, the largest known land animal that had ever lived (until several years later, with the discovery of the giant sauropod dinosaurs of the Morrison Formation. Cope and his team eventually recovered complete hip bones, sacral vertebrae, and several ribs from the animal. Since these were the first ceratopsian remains found, Cope was uncertain as to precisely what sort of dinosaur Agathaumas was (and for a time considered it a hadrosaur) until O. C. Marsh described Triceratops in 1889. In an 1889 paper, Cope suggested that Marsh's Ceratopsidae be renamed Agathaumidae, because of the paucity of Ceratops remains. Unfortunately, the bones of the rear half of the animal found are not particularly diagnostic in ceratopsians and Agathaumas remains a nomen dubium. No other remains have been found in the area, but based on its size and age of the rocks, it probably was a Triceratops. Хасмозаврины или цератопсины (Chasmosaurinae или Ceratopsinae) — подсемейство птицетазовых динозавров, обитавших в позднем меловом периоде (около 77—65 миллионов лет назад). Все известные виды были четвероногими и обитали в основном в Северной Америке хотя один вид, Тураноцератопс, известен из отложений Азии. Большинство представителей этого подсемейства обладают лишь шишковатыми «зачаточными» рожками, расположенными над глазницами, в отличие от своих ранних предков), тогда как трицератопс наоборот имеет крупные костяные наросты. Самым ранним из всех известных рогатых динозавров в Северной Америке является зуницератопс (Zuniceratops), живший 90 млн лет назад.
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, птицетазовые, хасмозаврины, цераподы, цератопсиды, цератопсы |
| 04:39 pm [industrialterro] [Link] |
Aerosteon Аэростеон (Aerosteon, от др.-греч. ἀερο- + ὀστέον «воздушная кость») — род ящеротазовых динозавров из семейства Neovenatoridae, подотряда тероподов, живший в меловом периоде около 85 — 74 миллионов лет назад, на территории нынешней Южной Америки. Окаменелости теропода были найдены в Аргентине. Описаны Полом Серено в 2008 году. Всего известен один скелет Aerosteon. Представлен единственным видом — A. riocoloradensis. В результате анализа дыхательной системы этого ящера было найдено сходство с современными птицами. Этот анализ окаменелостей показывает эволюционные связи между динозаврами и современными птицами. Aerosteon is a genus of either allosauroid or tyrannosauroid theropod dinosaur from the Late Cretaceous period of Argentina. Its remains were discovered in 1996 in the Anacleto Formation, dating to the Santonian stage (about 84 million years ago). The type and only known species is A. riocoloradense. Its specific name indicates that its remains were found 1 km north of the Río Colorado, in the province of Mendoza. They show evidence of a bird-like respiratory system. Aerosteon's name can be translated as air bone and derives from Greek ἀήρ (aer, "air") and ὀστέον (osteon, "bone"). Though the species name was originally published as "riocoloradensis", Greek ὀστέον is neuter gender, so according to the ICZN the species name must be riocoloradense to match. Aerosteon was a 9 metre (30 ft) long, 2 metric ton bipedal carnivorous dinosaur that lived approximately 83 million years ago during the Santonian stage. The remains discovered include a single tooth, some cranial bones, a number of partial or complete vertebrae from the neck, back, and sacrum, several cervical and dorsal ribs, gastralia, furcula (wishbone), left scapulocoracoid, left ilium, and left and right pubes. The incomplete fusion of some of its bones indicate that it was not quite fully mature. Aerosteon did not initially appear to belong to any of the three groups of large theropods that were known to have inhabited the southern continents during this time (namely the Abelisauridae, Carcharodontosauridae or Spinosauridae). Sereno suggested that it might be related to the allosauroid radiation of the Jurassic period, and this was confirmed in subsequent studies that recognized a clade of late-surviving, lightly built, advanced allosauroids with large hand claws similar to the spinosaurs, called the Megaraptora, within the allosaur family Neovenatoridae. A later analysis has placed Aerosteon within the Tyrannosauroidea. Some of Aerosteon's bones show pneumatisation (air-filled spaces), including pneumatic hollowing of the furcula and ilium, and pneumatisation of several gastralia, suggesting that it may have had a respiratory air-sac system similar to that of modern birds. These air sacs would have acted like bellows, moving air into and out of the animal's relatively inflexible lungs, instead of the lungs being expanded and contracted as occurs with mammals. See avian respiratory system for more detailed information on this. Sereno theorises that this respiratory system may have developed to assist with regulating body temperature and was later co-opted for breathing. Aerosteon was first described by Sereno et al. in a paper which appeared in the online journal PLoS ONE in September 2008. However, at the time, the International Code of Zoological Nomenclature did not recognize online publication of names for new species as valid unless print copies were also produced and distributed to several libraries, and that this action is noted in the paper itself. PLoS ONE initially failed to meet this requirement for Aerosteon. On 21 May 2009, the journal's managing editor coordinated with the ICZN to correct this oversight, publishing a comment to the original paper with an addendum stating that the requirements had been met as of that date. Consequently, though the description appeared in 2008, Aerosteon was not a valid name until 2009. Неовенаториды (Neovenatoridae) — семейство больших плотоядных динозавров, из надотряда Allosauroidea, живших в меловом периоде около 130—70 миллионов лет назад. Неовенаториды являются последними представителями аллозавроидов, по крайней мере один из их представителей — Orkoraptor жил в конце мелового периода (маастрихт), примерно 70 миллионов лет назад. Лопатки более короткие и широкие, маленькие, длинные передние конечности с большими когтями, приспособленные к быстрому бегу ноги и тазовые кости, которые для пневматизации имели заполненные воздухом полости и были легче по сравнению с другими Allosauroidea. Взрослые особи достигали размера от 175 кг (фукуираптор) до примерно 2500 кг (хилантаизавр). В 2009 году кладистический анализ проведенный Роджером Бенсоном и его коллегами, показал, что группа Carcharodontosauridae требует пересмотра, и необходимо было разделить ее на две ветви. Одна ветвь включала более традиционные виды, которые считались Carcharodontosauridae и сохранила свое название, другая ветвь состояла из Neovenator и некоторых родственных видов и была названа в 2010 году Neovenatoridae. Семейство Neovenatoridae было определено как: группа, состоящая из всех видов более тесно связаных с Neovenator salerii, чем Carcharodontosaurus saharicus, Allosaurus fragilis и Sinraptor dongi. ( Read More ) Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, карнозавры, неовенаториды, тероподы, ящеротазовые |
| 04:02 pm [industrialterro] [Link] |
Aeolosaurus Эолозавр (Aeolosaurus) — род травоядных динозавров из группы титанозавров инфраотряда зауроподов, живший в меловом периоде около 70—68 миллионов лет назад, на территории нынешней Южной Америки. Окаменелости четвероногого зауропода были найдены в Аргентине и Бразилии. Описаны Powell в 1987 году. Всего известно 3 вида эолозавров: Aeolosaurus colhuehuapensis. Aeolosaurus maximus — вид, описанный в 2011 году. Находка была сделана в формации Adamantina на территории современной Бразилии. Aeolosaurus rionegrinus. Aeolosaurus (/ˌiː.əlɵˈsɔrəs/; "Aeolus' lizard") is a genus of titanosaurian sauropod dinosaur from the Late Cretaceous Period of what is now South America. Like most sauropods, it would have been a quadrupedal herbivore with a long neck and tail. The remains of this dinosaur are incomplete, so size can only be estimated but Aeolosaurus was probably at least 45 feet (14 m) in length. This dinosaur is named after the Greek mythological figure Aeolus, Keeper of the Winds in Homer's Odyssey, because of the frequent winds that blow across Patagonia, where the remains were found. The generic name also includes the Greek sauros ('lizard'), the traditional suffix used in dinosaur names. The specific name (A. rionegrinus), refers to its location, in the Rio Negro Province of Argentina. Both genus and species were named and described by Argentine paleontologist Jaime Powell in 1987. The holotype of Aeolosaurus rionegrinus consists of a series of seven tail vertebrae, as well as parts of both forelimbs and the right hindlimb. It was discovered in the Angostura Colorada Formation in Argentina, which dates from the Campanian stage of the Late Cretaceous, about 83 to 74 million years ago. Relationships among the many titanosaurian sauropods are hazy at best, but Aeolosaurus has been tentatively linked to a few other genera, based on features of the tail vertebrae, including Rinconsaurus and Adamantisaurus. Gondwanatitan and Aeolosaurus both exhibit neural spines on the tail vertebrae that point forwards, a feature not seen in any other known titanosaurians. The vertebrae from the middle part of its tail had elongated centra. Aeolosaurus had vertebral lateral fossae that resembled shallow depressions. Fossae that similarly resemble shallow depressions are known from Saltasaurus, Alamosaurus, Malawisaurus, and Gondwanatitan. Venenosaurus also had depression-like fossae, but its "depressions" penetrated deeper into the vertebrae, were divided into two chambers, and extend farther into the vertebral columns. Its middle tail vertebrae's neural spines angled anteriorly when the vertebrae are aligned. These vertebrae resemble those of Cedarosaurus, Venenosaurus, and Gondwanatitan. Another specimen, described in 1993, consists of five tail vertebrae, and some bones from the forelimb and pelvis. Since there are two right ulna (forearm) bones, the specimen must consist of at least two individuals. These bones were also associated with two osteoderms, or bony armor plates, providing evidence that this dinosaur was armored. This specimen is from the Allen Formation of Rio Negro, dating back about 70 to 68 million years ago to the middle Maastrichtian stage. While this specimen bears features that characterize the genus Aeolosaurus, it is from a younger time period and shows enough differences that the authors recognized it as a possible second species. Another partial skeleton, including four more tail vertebrae and material from both limbs on the left side of the body, was described in 1997. This was recovered from the Los Alamitos Formation of Rio Negro, which falls between the other two dates. This specimen was also referred to the genus Aeolosaurus, but not to the species A. rionegrinus, and may represent a third species. However, since the genus Aeolosaurus is not well known, the authors chose not to formally name either of these possible new species. For now, they are both simply known as "Aeolosaurus sp." Future discoveries may give scientists more information on variation within the genus, and show that all of the above specimens belong to A. rionegrinus, or that they merit being formally named. Another series of 15 tail vertebrae was assigned to Aeolosaurus in the original description, but it was later determined that the series does not belong to this genus, as it lacks several features found in the other specimens of Aeolosaurus. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| February 17th, 2015 | |
| 09:54 pm [industrialterro] [Link] |
Aegyptosaurus Египтозавр (Aegyptosaurus; букв. «ящер из Египта») — род травоядных динозавров из группы титанозавров, инфраотряда зауроподов, живший в конце мелового периода (альб и сеноман) около 95 миллионов лет назад, на территории нынешней Африки. Окаменелости четвероногого зауропода были найдены в Египте, Нигере и в различных местах пустыни Сахара. Вид описан Штормером в 1932 году. Всего известно около семи скелетов Aegyptosaurus. Соотношение костей плечевой кости к бедру - 0,78, локтевой к плечевой - 0,75, голени к бедру - 0,69. Всего известно около семи скелетов египтозавра. Египтозавры по строению тела очень похожи на гигантских динозавров - представителей титанозавридов - аргентинозавров. Вполне возможно, египтозавры являются потомками аргентонозавров. Это предположение основано также на гипотезе о существовании когда-то «моста» между континентами, по которому некоторые виды древних животных расселялись по всей суше. Aegyptosaurus /iːˌdʒɪptɵˈsɔrəs/ meaning 'Egypt’s lizard', for the country in which it was discovered (Greek sauros meaning 'lizard') is a genus of sauropod dinosaur believed to have lived in what is now Africa, around 95 million years ago, during the mid- and late-Cretaceous Period (Albian to Cenomanian stages). Like most sauropods, it had a long neck and a small skull. The animal's long tail probably acted as a counterweight to its body mass. Aegyptosaurus was a close relative of Argentinosaurus, a much larger dinosaur found in South America. Aegyptosaurus was described by German paleontologist Ernst Stromer in 1932. Its fossils have been found in Egypt, Niger and in several different locations in the Sahara Desert. All known examples were discovered before 1939. The fossils were stored together in Munich, but were obliterated when an Allied bombing raid destroyed the museum where they were kept in 1944, during World War II. It is possible that Aegyptosaurus was common prey for large predatory dinosaurs, such as Carcharodontosaurus and Spinosaurus. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| February 15th, 2015 | |
| 07:36 pm [industrialterro] [Link] |
Adasaurus Адазавр (Adasaurus; буквально «ящерица Ады») — род тероподовых динозавров, из надсемейства дромеозавридов, живших в позднем меловом периоде, на территории нынешней Центральной Азии. Это был маленький, двуногий хищник со серповидными когтями на втором пальце каждой задней ноги. В размере соизмерим со взрослым человеком, вероятно, около 2 метров, от головы до кончика хвоста. Назван в честь Ады, злого духа в национальной мифологии Монголии. Имя также включает в себя др.-греч. σαῦρος — «ящерица», наиболее распространенное слово, используемое в именах динозавров. Существует один вид (А. mongoliensis), названный в честь страны Монголии, где ископаемые были найдены. Род и вид были названы и описаны в 1983 году известным монгольским палеонтологом Р. Барсболдом. Adasaurus is a member of Dromaeosauridae, a group that is closely related to living birds. Other dromaeosaurids include Deinonychus, Velociraptor, Microraptor, and Buitreraptor. The relationships of Adasaurus are poorly understood. Traditionally, Adasaurus is assigned to the Dromaeosaurinae, which includes heavily built animals such as Dromaeosaurus and Utahraptor but several recent studies have suggested that it may be a member of the Velociraptorinae instead. Two specimens of Adasaurus have been found, both from the Nemegt Formation in the Gobi Desert of southern Mongolia. The holotype, IGM 100/20, is an incomplete skeleton with partial skull, including the vertebral column except the back of the tail, all three bones of the pelvis, the shoulder girdle and the hindlimbs. The second specimen, the paratype IGM 100/51 also described in the original paper, consists of the back end of another skeleton, including the hindlimbs. Both specimens are currently in the collection of the Mongolian Geological Institute in Ulaanbaatar, Mongolia. The age of the Nemegt is not known for certain, but it is commonly thought to belong to the Maastrichtian stage of the Late Cretaceous Period., and Adasaurus would therefore have lived between 72 and 66 million years ago. Other dinosaurs found in this formation include the tyrannosaur Tarbosaurus, the ornithomimid Anserimimus, the troodontid Zanabazar, and the hadrosaur Saurolophus. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, дейнонихозавры, диапсиды, динозавроморфы, динозавры, дромеозавриды, манирапторы, тероподы, целурозавры, ящеротазовые |
| 06:12 pm [industrialterro] [Link] |
Adamantisaurus Адамантизавр (Adamantisaurus) — род динозавров из группы титанозавров, инфраотряда завроподов, живших в позднем мелового периоде, на территории современной Южной Америки. Известен только по шести хвостовым позвонкам, но, так как этот ящер является завроподом, можно предположить, что этот динозавр был очень крупным животным с длинной шеей и хвостом. Точный внешний вид восстановить невозможно, пока не будет найден более полный материал. Как и все завроподы, это был крупный, четырехногий травоядный динозавр с длинными шеей и хвостом. До сих пор науке известны лишь 6 соединенных друг с другом передних хвостовых позвонков, а также 2 нижних гемапофиза (отростки хвостовых позвонков), которые были обнаружены в бразильском штате Сан-Паулу. Род Адамантизавр, представленный единственным видом Adamantisaurus mezzalirai, впервые был описан в 2006 году бразильскими палеонтологами Сантуччи (Santucci) и Бертини (Bertini). Родовое название «Adamantisaurus» указывает на принадлежность ископаемых останков динозавра к формации Адамантина (Adamantina). Видовое название «mezzalirai» дано в честь палеонтолога Серджио Меццалиры (Mezzalira), собравшего кости и оставившего первое упоминание о них в научной литературе. Таким образом, полное название «Adamantisaurus mezzalirai» может быть переведено на русский язык как «Адамантизавр Меццалиры». В момент обнаружения 6 хвостовых позвонков были анатомически связаны между собой. Предполагается, что это – ряд хвостовых позвонков со второго по седьмой. Позвонки сильно вогнуты с передней стороны. Они имеют несколько автапоморфий (отличительных признаков), позволяющих отделить этот род от других: так, например, направленные вверх отростки позвонков чуть отогнуты назад и имеют увеличенные с боков верхние концы; постзигапофизы и презигапофизы (механические элементы соединения позвонков) тоже имеют увеличенные поверхности сочленения, причем поверхность сочленения постзигапофизов вогнута. Эти позвонки еще в 1958 году, во время строительства железной дороги из Адамантины в Ирапуру, обнаружил палеонтолог Серджио Меццалира (Sergio Mezzalira) вместе с несколькими зубами титанозавров. Место находки расположено близ города Флорида-Паулиста на юго-западе штата Сан-Паулу и относится к геологической формации Адамантина (Adamantina formation), входящей в свиту пластов Бауру (Bauru). Точный возраст свиты Бауру до сих пор оспаривается, однако авторы первого описания адамантизавра предполагают, что кости относятся к слоям от позднего кампанского яруса до раннего маастрихта. Первоначально (в 1966, 1989 годах) Меццалира приписал одной и той же особи адамантизавра не только обнаруженные позвонки, но и бедренную кость, извлеченную строителями железной дороги из того же места, где позже нашлись и позвонки. Однако авторы первого научного описания посчитали, что эта кость принадлежала, вероятно, не этому адамантизавру, а другой особи, так как она, например, относительно мала по сравнению с позвонками и отличается от них по степени сохранности. Кроме того, в формации Адамантина часто встречаются места, в которых рядом обнаруживается сразу несколько видов, что позволяет предположить, что упомянутая бедренная кость могла принадлежать другому виду. Титанозавры (Titanosauria) — группа ящеротазовых динозавров инфраотряда зауроподов, живших в юрском и меловом периодах около 171—65,5 миллионов лет назад на всех континентах планеты, кроме Антарктиды. Являлись травоядными динозаврами, обладавшими длинными шеями и хвостами, и передвигавшиеся на четырёх ногах. В длину достигали до 35—40 метров и весили между 88 и 110 тоннами. ( Read More )
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, титанозавры, ящеротазовые |
| February 11th, 2015 | |
| 05:12 pm [industrialterro] [Link] |
Acrocanthosaurus Акрокантозавр (Acrocanthosaurus, также Acracanthus) — гигантский хищный динозавр раннего мелового периода, живший около 125—112 миллионов лет назад. Принадлежит к семейству Carcharodontosauridae, близкому к аллозаврам. Один из немногих кархародонтозаврид, найденных вне южных материков. Описан в 1950 году из раннего мела Оклахомы. Первоначально считался спинозавром из-за удлиненных остистых отростков. В настоящее время его относят либо к настоящим аллозаврам (Allosauridae) либо к кархародонтозаврам. Это огромное животное, с черепом длиной около 120 см, общая длина доходила до 13 метров. Череп сходен по форме с черепом кархародонтозавра и аллозавра — низкий, с орнаментацией на верхней поверхности. Остистые отростки достигали 35 см в высоту, образуя подобие низкого «паруса». В то же время, в отличие от паруса диметродона, остистые отростки «парусных» динозавров (спинозавра, уранозавра) очень толстые. Для акрокантозавра предполагается связь высоких остистых отростков с развитием очень мощных спинных мышц — таким образом, при жизни «паруса» как такового могло и не быть. Ноги относительно короткие, передние лапы также короткие, массивные, с очень крупными когтями (особенно на первом пальце). Шея мощная, прямая, морда при ходьбе была опущена примерно под углом 25° к грунту. Следы, возможно, принадлежащие акрокантозавру, обнаружены в раннемеловых отложениях Пэлакси в Техасе вместе со следами завропод. Судя по следам, хищник преследовал стадо. Акрокантозавр вполне мог атаковать и взрослого завропода средних размеров, особенно если жертва была больна или ослаблена. Не исключено, что акрокантозавры охотились стаями. Многочисленные остатки акрокантозавров (включая полные скелеты) известны из апта-альба Оклахомы, Аризоны, Техаса, Юты, Мэриленда. Это был верховный хищник так называемой формации Кловерли. Хотя он немного меньше, чем его гигантский родственники, Акрокантозавр был одним из крупнейших тероподов, когда-либо существовавших на земле. Крупнейший известный экземпляр (NCSM 14345), по оценкам, составляет 11,5 метров от морды до кончика хвоста и весил от 5,7 до 6,2 тонн, хотя некоторые ученые отмечают, что некоторые Акрокантозавры могли достигать и 7 тонн. Его череп был около 1,3 метров в длину. Череп Акрокантозавра, как и у большинства других аллозавров, был вытянутым, удлиненным и прямым. Этот динозавр мог охотится практически на всех травоядных динозавров своей экосистемы. При и без того громоздком теле, ему еще более впечатляющий вид придавали отростки на спинных позвонках, вместе образующие горб или парус, служивший или для отпугивания врагов и привлечения партнеров или для терморегуляции. Развивал скорость до 40 км/ч. В пасти акрокантозавра насчитывалось 86 острых зубов, которыми он без труда отрывал мясо жертвы. Если зуб обламывался, то на его месте вырастал другой. Когти росли на трехпалых передних конечностях, по форме напоминали косу, которыми он вцеплялся в добычу и удерживал ее. Тяжелый, низкий череп с характерным рельефным рисунком на поверхности головы чем-то напоминал череп других карнозавров: аллозавра и кархародонтозавра. Роговые, 35-ти сантиметровые отростки образовывали на голове целый частокол толстых наростов, что сильно отличало акрокантозавра от представителей «парусных» доисторических монстров: уранозавра и спинозавра. Благодаря сильным развитым мышцам спины эти отростки и были такими развитыми. Этого динозавра отличали мощная, прямая морда и сильные ноги. ( Read More ) Не классифицированные ископаемые останки представителей семейства известны с поздней юры (титонский век, 150,8—145,5 млн лет назад). Одним из самых ранних известных науке кархародонтозавридов является открытый в 2010 году Concavenator, живший около 130 миллионов лет назад. Наряду со спинозавридами, кархародонтозавриды были крупнейшими хищниками в раннем и среднем мелу, жившими по всей Гондване, с разновидностями также населявших территорию нынешних Северной Америки (Acrocanthosaurus и Siats meekokorum) и Азии (Shaochilong). Период их жизни колеблется от баррема (127—121 миллион лет назад) до турона (93-89 миллионов лет назад). Однако, после турона нет никаких свидетельств присутствия этих животных где-либо. Их место заняли меньшие в размерах абелизавриды в Гондване и тираннозавриды в Северной Америке и Азии. Согласно Фернандо Новасу и его коллегам, исчезновение не только кархародонтозаврид, но также и спинозаврид и других видов в Гондване и в Северной Америке указывает, что эта фаунистические изменения принимали глобальные масштабы. Семейство было выделено в 1931 году немецким палеонтологом Ernst Freiherr Stromer von Reichenbach (12.10.1870 — 18.12.1952) для единственного тогда нового вида Carcharodontosaurus saharicus и других крупнейших хищников планеты. Многие палеонтологи включают сюда род Acrocanthosaurus (Sereno et al. 1996, Harris 1998, Holtz 2000, Rauhut 2003), хотя другие учёные помещают его в близкое семейство Allosauridae (Currie & Carpenter, 2000; Coria & Currie, 2002). Род Neovenator также включен в состав Carcharodontosauridae (Rauhut, 2003; Holtz et al., 2004). После открытия Mapusaurus в 2006 году палеонтологи Rodolfo Coria и Phil Currie создали подсемейство Giganotosaurinae, включив в него многие виды Южной Америки, отличающиеся от Африканских и Европейских форм. К этому подсемейству также может быть отнесён и род Tyrannotitan, описание которого пока не позволяет сделать однозначные таксономические выводы. Репродукции (1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12): ( Read More ) Размеры тела в сравнении с человеком:
Ископаемые останки (1, 2, 3, 4, 5): Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, карнозавры, кархародонтозавриды, тероподы, ящеротазовые |
| January 21st, 2015 | |
| 05:18 pm [industrialterro] [Link] |
Achillobator Ахиллобатор (Achillobator) — род динозавров, принадлежавший семейству дромеозавриды. Ахиллобатор обитал на территории современной Монголии около 90 миллионов лет назад. Известен только один вид данного рода - Achillobator giganticus. Название этого динозавра состоит из двух слов - имени героя Троянской войны Ахилла и монгольского слова "bator", означающего "воин" или "герой". Видовое название "giganticus" было присвоено этому ящеру благодаря тому, что ахиллобатор значительно превосходит размерами большинство дромеозаврид. Длина ахиллобатара составляла примерно от 4,6 до 6 метров, что делает его вторым по величине в размерном классе среди дромеозаврид после ютараптора. Как уже отмечалось выше, ахиллобатор принадлежал к семейству дромеозаврид. Согласно последним исследованиям, ахиллобатор находится в наиболее тесном родстве с североамериканскими дромеозавридами, такими как ютараптор и дромеозавр, вместе с которыми он представляет подсемейство дромеозаврин (Dromaeosaurinae). Судя по всему, таз ахиллобатора показывает некоторые примитивные черты ящеротазовых динозавров (Saurischia) в сравнении с другими дромеозавридами. Например, лонная кость поставлена вертикально и имеет большой "ботинок" (расширение на конце) в отличие от большинства других дромеозаврид, у которых "ботинок" намного меньше, если таковой вообще имеется, и лонная кость которых направлена назад в том же направлении, что и седалищная кость (состояние, называемое опистопубией, независимое появление которой отмечено у теризинозавров и птицетазовых, а также у птиц и их ближайших родственников). Впервые, останки ахиллобатора были обнаружены во время советско-монгольской экспедиции в 1989-ом году, однако вид был описан только 10 лет спустя - в 1999-ом году монгольским палеонтологом Алтангерелем Перле (Altangerel Perle) и американцами Марком Нореллом (Mark Norell) и Джимом Кларком (Jim Clark), хотя описание было не полным и издавалось фактически без участия двух последних авторов. Окаменелые кости ахиллобатора были в основном разъединёнными, но связанными между собой, среди которых были фрагмент верхней челюсти с зубами, позвонки всех отделов позвоночника, рёбра, плечевые кости, кости таза и кости передних и задних конечностей. Эти останки были найдены в Bayan Shireh Formation в провинции Dornogovi, Монголия и были датированы поздним меловым периодом. Achillobator (/əˌkɪləˈbaɪtɔr/ a-KILL-ə-BAY-tohr) is a dromaeosaurid theropod dinosaur that lived roughly 98 to 83 million years ago during the Late Cretaceous in what is now Mongolia, in Asia. It was among the largest dromaeosaurs; the holotype and only known individual of Achillobator is estimated at 5 to 6 m (16.4 to 19.7 ft) long. Achillobator was a moderately-built, ground-dwelling, bipedal carnivore. It would have been an active predator, hunting with the enlarged, sickle-shaped claw on the second toe. The fossils of the type specimen of Achillobator, FR.MNUFR 15, were found associated but mostly disarticulated, and include a left maxilla with teeth, two cervical vertebrae, two dorsal vertebrae, rib fragments, seven caudal vertebrae, a scapula and coracoid, a pelvis with a right ilium, pubis and ischium, a radius, an incomplete manus, a left femur and tibia, and an incomplete pes. Smith at al. (2012) noted that this genus represents the second largest of the known dromaeosaurid taxa with a tibial length of 490 mm. Its femur, which is 3% longer than the tibia, a rare trait in Dromaeosaurs, measures 505 mm in length. Estimates suggest that Achillobator weighed 350 kilograms (771.6 pounds) at most. The teeth are serrated and recurved, and the posterior serrations are slightly larger than the anterior serrations. The pelvis of Achillobator seems to show plesiomorphic ("primitive") saurischian characteristics compared to other dromaeosaurids. For instance, the pubis is aligned vertically and has a large pubic boot (a wide expansion at the end), unlike most other dromaeosaurids, where there is a much smaller boot, if any, and the pubis points backwards in the same direction as the ischium (a condition called opisthopuby, which is also seen in the unrelated therizinosaurs and ornithischians, as well as in birds). The above differences and others led Burnham et al. (2000) to suggest that Achillobator represents a paleontological chimera. However, other studies have attempted to refute this, noting that many pieces were found to be semi-articulated, all of the elements are the same color and preservation, and that Achillobator routinely comes out as a dromaeosaurid in cladistic analyses, even taking into account the differences. Achillobator is a dromaeosaurid, a family of dinosaurs currently thought to be very closely related to birds. While the relationship of dromaeosaurids to other theropods (including birds) is relatively well-understood, the phylogeny within the family itself is not. The more recent phylogenetic analyses, conducted by Longrich and Currie (2009) and Senter et al. (2012) show that Achillobator is a member of the subfamily Dromaeosaurinae, most closely related to North American forms like Utahraptor and Dromaeosaurus. Deinonychus and Velociraptor are also dromaeosaurids, but appear to represent a different branch of the family. A diagnosis is a statement of the anatomical features of an organism (or group) that collectively distinguish it from all other organisms. Some, but not all, of the features in a diagnosis are also autapomorphies. An autapomorphy is a distinctive anatomical feature that is unique to a given organism or group. According to the revised diagnosis by Turner et al. (2012), Achillobator can be distinguished based on the following combination of characteristics and autapomorphies:
The remains of Achillobator were recovered in the Bayan Shireh Formation of Dornogovi Province, Mongolia, in fine-grained, medium sandstone/gray mudstone that was deposited during the Late Cretaceous epoch. The exact age is uncertain, with two competing hypotheses; based on comparisons with other formations, the Bayan Shireh fauna seems to correspond best with the Turonian through early Campanian stages of the Late Cretaceous, about 93 to 80 million years ago. However, examination of the magnetostratigraphy of the formation seems to confirm that the entire Bayan Shireh lies within the Cretaceous Long Normal, which lasted only until the end of the Santonian stage, giving a possible Cenomanian through Santonian age, or between 98 and 83 million years ago. The paleofauna of the Bayan Shireh Formation included the tyrannosaurid Alectrosaurus, the therizinosaurid Segnosaurus, the ankylosaurid Talarurus, the hadrosaurid Bactrosaurus, as well as a genus of sauropod, and an azhdarchid pterosaur. Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, дейнонихозавры, диапсиды, динозавроморфы, динозавры, дромеозавриды, манирапторы, тероподы, целурозавры, ящеротазовые |
| December 30th, 2014 | |
| 08:35 pm [industrialterro] [Link] |
Achillesaurus Ахиллезавр (Achillesaurus) — род ящеротазовых динозавров семейства альваресзавриды, живших в меловом периоде около 85—83 млн лет назад на территории нынешней Аргентины. Был описан в 2007 году Агустином Мартинелли и Эсекьелем Верой на основе частичного скелета, состоящего из крестцового позвонка, четырёх костей хвостовых позвонков, части левой бедренной кости, голени и стопы, а также левой подвздошной кости. Исследователи отнесли новый род к семейству альваресзавриды с неясным отношением к другому динозавру этого семейства — Alvarezsaurus. Achillesaurus is a genus of alvarezsaurid theropod dinosaur from the Santonian-age Upper Cretaceous Bajo de la Carpa Formation of Rio Negro, Argentina. It was named in reference to Achilles' heel, because diagnostic features are found there for these animals. The genus was a relatively large, basal alvarezsaurid, and a contemporary of Alvarezsaurus. Achillesaurus is based on MACN-PV-RN 1116, a partial skeleton including a sacral vertebra, four tail vertebrae, part of the left thighbone, shin and foot, and the left ilium. Agustín Martinelli and Ezequiel Vera, who described the specimen, performed a phylogenetic analysis and found their new genus to be an alvarezsaurid with an unresolved relationship to Alvarezsaurus and more derived alvarezsaurids. Makovicky, Apesteguía & Gianechini (2012) argued that Achillesaurus might actually be a junior synonym of Alvarezsaurus which, according to the authors, "is known from the same formation and from which it [i.e. Achillesaurus] differs trivially". Альваресзавриды (Alvarezsauridae) — небольшое семейство мелких тероподов, обладающих признаками, как птиц, так и динозавров. Жили 160—70 миллионов лет назад. Изначально альваресзавриды считались ранними птицами (важные признаки птиц у альваресзавридов — оперение и теплокровность), но сейчас многие учёные считают их примитивными манирапторами. Есть и третья версия, согласно которой альваресзавриды - это близкие родственники орнитомимозавров (Ornithomimosauria). Названы в честь историка Дона Грегорио Альвареса. Самые ранние родственники альваресзаврид согласно последним археологическим данным обитали 160 млн лет назад (динозавр Haplocheirus) на территории современного Западного Китая, из-за чего учёные предполагают, что представители этой клады изначально эволюционировали в Азии. Изучив все особенности данного динозавра, учёные сделали вывод, что альваресзавриды эволюционировали параллельно с птицами. Новый динозавр подтверждает, что надсемейство Alvarezsauroidea занимает базальное положение в группе Maniraptora. Род Haplocheirus также является одним из крупнейших представителей альваресзаврид. А это означает, что Alvarezsauridae постепенно уменьшались в размере в ходе эволюции. ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, альваресзавриды, альваресзавры, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, манирапторы, тероподы, целурозавры, ящеротазовые |
| December 28th, 2014 | |
| 09:24 pm [industrialterro] [Link] |
Achelousaurus Ахелоузавр (Achelousaurus — «ящер Ахелоя») — род динозавров из группы цератопсов, живших в позднем меловом периоде 75—70 млн лет назад (с кампанского по маастрихский ярусы позднего мела), на территории нынешней Монтаны (США). Представлял собой четвероногого травоядного динозавра с попугаеобразным клювом, небольшими костными шишками на морде и двумя над глазницами, а также двумя рогами на вершине костистого нароста на шее. По мере роста животного костный нарост на носу увеличивался и становился направленным вперёд, а наросты над глазами становились бугристыми. Ахелоузавр достигал длины 6 метров, высоты 3,5 метра и веса в 2,5 тонны. Череп ахелоузавра достигал 1,6 метра в длину. В 1995 году палеонтолог Скотт Сэмпсон описал единственный вид этого рода — A. horneri. Видовой эпитет дан в честь Джека Хорнера, американского палеонтолога, известного своими открытиями динозавров в штате Монтана. Родовое название Achelousaurus дано в честь персонажа греческой мифологии Ахелоя — божества, в борьбе с Гераклом потерявшего один из рогов. На данный момент найдено всего три скелета ахелоузавров. The genus and the one named species (A. horneri) were both named by paleontologist Scott Sampson in 1995. The specific name honors Jack Horner, an influential American paleontologist famous for his Montana dinosaur discoveries, who in 1987 headed the team that excavated the holotype skull of Achelousaurus, MOR 485. The generic name Achelousaurus is a complex reference to Greek mythology. Achelous, an important Greek river deity, had one of his horns torn off by Hercules, in a mythological fight with the legendary hero. All three known skulls of Achelousaurus have rough bosses in the same places where other ceratopsians have horns, giving them the appearance of having had the horns ripped off. Achelous was also celebrated for his shapeshifting ability, just as Achelousaurus appear to combine features of other ceratopsian dinosaurs. Achelousaurus horneri are known from the U.S. state of Montana, in the Two Medicine Formation, which preserves sediments dated from the Campanian stage of the Late Cretaceous Period, between 83 and 74 million years ago. Achelousaurus specimens are found in the highest levels of the formation, probably closer to the end of that timeframe, 74 mya. Contemporary dinosaur species included Prosaurolophus blackfeetensis, Scolosaurus cutleri, Hypacrosaurus stebingeri, Einiosaurus procurvicornis, and tyrannosaurids of uncertain classification. Scientists have so far recovered three skulls and some postcranial material from the Two Medicine, all housed at the Museum of the Rockies in Bozeman, Montana. The skull of a full-grown Achelousaurus (including the frill horns) is over 5 feet (1.6 meters) long. Early reports suggested that Achelousaurus represented a transitional form, "Type C", between ceratopsians with modified horns like Einiosaurus, "Type B" (with which A. horneri shares two horns on the end of the frill), and the derived, hornless Pachyrhinosaurus (Horner et al., 1992). While they may or may not form a direct line of descent, all three of these genera are at least closely related, and are often united in the tribe Pachyrhinosaurini, inside the subfamily Centrosaurinae and the family Ceratopsidae (Sampson, 1995; Dodson et al., 2004). ( Read More ) Размеры тела в сравнении с человеком:
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, маргиноцефалии, неорнитискии, птицетазовые, центрозаврины, цераподы, цератопсиды, цератопсы |
| December 23rd, 2014 | |
| 09:20 pm [industrialterro] [Link] |
Abydosaurus Абидоза́вр (Abydosaurus; букв. «ящер Абидоса») — род ящеротазовых динозавров из семейства брахиозавриды инфраотряда зауропод, живших в раннем меловом периоде (около 105—104 млн лет назад) на территории Северной Америки (ныне штаты Юта и Колорадо, США). Представлен единственным видом — Abydosaurus mcintoshi. Первые останки динозавра, принадлежащему этому роду, были обнаружены в США на дне карьера на территории национального парка Dinosaur National Monument, на границе штатов Юта и Колорадо. В 2009 году для извлечения костей из горных пород костей динозавров на территории парка был использован динамит. Описан в 2010 году группой палеонтологов под руководством Daniel Chure. Всего палеонтологи нашли разрозненные кости скелетов четырёх молодых особей динозавров и четыре черепа, два из которых сохранились практически полностью. Длина молодых особей этого вида составляла 7-8 метров, размер взрослых представителей был, вероятно, гораздо крупнее. Название «абидозавр» является ссылкой на египетскую мифологию — Абидос в Древнем Египте был основным центром поклонения Осирису; как считалось, здесь захоронена голова бога (тогда как голотип абидозавра состоял из черепа и шеи). Данный типовой вид был назван Abydosaurus mcintoshi в честь палеонтолога Джека Макинтоша (John S. ("Jack") McIntosh). Абидозавр жил приблизительно 105 миллионов лет назад, в конце раннего мелового периода, в альбском веке. Исследование костей Abydosaurus показали его тесное родство и, вероятно, происхождение от более раннего завропода брахиозавра. В связи с этим D. J. Chure (2010) выделяет данный род в семейство Brachiosauridae. It is of interest because it is one of the few sauropods known from skull material, with the first described complete skull for a Cretaceous sauropod from the Americas. It is also notable for its narrow teeth, as earlier brachiosaurids had broader teeth. Abydosaurus is based on DINO 16488, a nearly complete skull and lower jaws with the first four neck vertebrae. Abundant skull and postcranial bones were found at the same site, including partial skulls from three additional individuals, a partial hip and associated tail vertebrae, a shoulder blade, an upper arm bone, and hand bones. These fossils were found in a sandstone bed at the base of the Mussentuchit Member of the Cedar Mountain Formation, near the old visitor center of Dinosaur National Monument. Zircons from mudstones beneath the bone-bearing sandstone indicate the age of the sandstone and the its contained bones is less than 104.46 ± 0.95 million years, in the Albian stage of the Early Cretaceous. Although Abydosaurus lived some 45 million years after Giraffatitan, - the skulls of these two genera are similar except for the narrower, sharper teeth and smaller nose of Abydosaurus. Abydosaurus can be differentiated from all other sauropods, including Giraffatitan, by subtle features of the nasal and maxillary bones, its relatively small external nares (nostrils), and some features of the teeth.
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, брахиозавриды, диапсиды, динозавроморфы, динозавры, завроподоморфы, завроподы, макронары, титанозавроформы, ящеротазовые |
| December 14th, 2014 | |
| 07:05 pm [industrialterro] [Link] |
Abelisaurus Абелизавр (Abelisaurus — «ящер Абеля») — род плотоядных динозавров из семейства абелизаврид, подсемейства абелизавров, обитавший 84-80 млн лет назад. Останки абелизавра были обнаружены в провинции Неукен (Аргентина). Скелеты абелизаврид были найдены на всех южных континентах, кроме Антарктиды. Свое имя они получили в честь Роберто Абеля (Roberto Abel), директора аргентинского музея (Museo Provincial de Cipolletti Carlos Ameghino), в котором были выставлены останки абелизавра. Абелизавры были крупными тероподами, с длиной тела до 9 метров, высотой в 3,5 метра и весом до 2,5 тонн. У них был высокий череп с широкой межорбитальной крышей и кинжалообразными зубами. Передние конечности — небольшие, с острыми когтями, расположены высоко на теле. Задние конечности — мощные и мускулистые. Очевидно, что абелизавры могли развивать большую скорость, были подвижными и быстрыми. Абелизавры были хищниками, охотясь на других динозавров. Абелизавр обладал исключительными свойствами строения: в высоком черепе абелизавра, длинном, как крикетная бита (от восьмидесяти пяти сантиметров до одного метра) с широкой межорбитальной крышей прямо над челюстями, имелось большое отверстие. Оно уменьшало массу головы, благодаря чему абелизавр мог быстро перемещаться, был подвижным и быстрым. Как и у тираннозавра (который, как утверждают палеонтологи, возможно приходился ему дальним родственником), небольшие передние конечности с острыми когтями располагались высоко на туловище абелизавра. С их помощью абелизавр рвал на куски мясо добычи. Лапы животного имели по четыре пальца. Каждый коготь заканчивался острым наконечником, что позволяло хищнику крепко впиваться в плоть несчастной жертвы. В течение жизни когти покрывались оболочкой из плотного рогового вещества, называемого кератином. Этот же материал лежит в основе наших волос и ногтей. По мере того как оболочка истиралась, формировался очень острый кончик когтя, образуя тем самым эффективный инструмент позволяющий резать и рвать. Абелизавр был страшным и грозным хищником. Устрашающий вид этим древним ящерам также придавали кинжалоподобные зубы. Пасть абелизавра была вооружена множеством острых, загнутых внутрь зубов, на верхней челюсти в ее передней части было около тринадцати зубов. Эти зубы идеально подходили для раздирания сырого мяса. Запустив в тело жертвы свои острые кривые зубы, абелизавр вырывал куски мяса резко запрокидывая голову. Жертва, стиснутая гигантскими челюстями абелизавра, практически не имела шансов на спасение. На верхней части морды с крючковатым носом, по носовой и предлобной костям, вероятно, находились бугристости, чье предназначение остается неясным. Этот ящер получил свое имя «Abelisaurus» в честь директора аргентинского провинциального музея Ципполети (Museo Provincial de Cipolletti Carlos Ameghino) Роберто Абеля (Roberto Abel), который занимался изучением и описанием этого вида динозавров. Позднее скелет абелизавра был включен в экспозицию музея и с разрешения Роберто Абеля впервые был выставлен на всеобщее обозрение публике. Первоначально абелизавр был изначально описан из формации Аллен, но последующее исследование доказало, что останки были фактически найдены в формации Анаклето (часть группы Неукен) провинции Рио-Негро, Аргентина. Анаклето - геологическая формация в Южной Америке, датируемая ранним кампаном позднего мелового периода, между восьмидесятью тремя и восьмидесятью миллионами лет назад. К сожаленью скелет был найден далеко не в полном виде, был обнаружен только череп, по большей части его левая сторона, также отсутствовало большинство костей неба. Однако, даже по такой небольшой находке можно получить представление о размере динозавра и его внешнем облике. Изучая череп животного ученые обнаружили некоторое сходства с другим аргентинским тероподом - карнотавром. Это открытие ученых – палеонтологов указывало на существование неизвестной группы плотоядных ящеров-теропод, бродящих по Аргентине во времена мелового периода примерно в то время, когда тиранозавр жил в северном полушарии. Карнотавр и абелизавр были так необычны и так не похожи на другие семейства, что ученые решили их выделить в отдельное семейство в полностью новую группу динозавров – абелизавриды. Это семейство отделилось от остальных теропод в средней юре, примерно 170 млн. лет назад. После открытия абелизавра было найдено много других абелизаврид, включая практически полностью сохранившиеся скелеты «Aucasaurus», «Carnotaurus» и «Majungasaurus». Некоторые палеонтологи помещают абелизавра как примитивного абелизаврида, в состав подсемейства «Abelisaurinae», а все остальные таксоны, за исключением неопределенных, размещены в подсемейство «Carnotaurinae». Другие палеонтологи менее уверены относительно его систематического положения. Абелизавриды разделяют некоторые особенности строения черепа с другими крупными тероподами из семейств кархародонтозаврид и тираннозаврид, однако, это сходство по большей части внешнее. Предковой формой абелизавра и всех его родственников с достаточной долей уверенности можно считать среднеюрских цератозаврид. В 1996 году Жозе Бонапарте отметил, что носовые кости абелизавра с сильно морщинистой поверхностью (кроме участка у носового отверстия) имеют очень грубое покрытие. Также он заметил, что морда у абелизавра очень высокая и узкая, а это является прямым свидетельством того, что абелизавр был жестоким хищником. Другие признаки того, что абелизавр был активным хищником – большие надвисочные и предглазничные отверстия (очевидно, для размещения больших мускулов челюсти), а также острые, суженные поперек и с короткими коронами зубы. ( Read More ) Размеры тела в сравнении с человеком: Tags: Вымершие рептилии, Мел, абелизавриды, абелизавры, авеметатарзалии, архозавроморфы, архозавры, диапсиды, динозавроморфы, динозавры, тероподы, цератозавры, ящеротазовые |
| November 30th, 2014 | |
| 11:17 pm [industrialterro] [Link] |
Vectidraco Vectidraco daisymorrisae — вид птерозавров из надсемейства Azhdarchoidea. Обнаружен в породах мелового периода (Atherfield Clay Formation, аптский ярус) на острове Уайт (Южная Англия, Великобритания). Возраст находки — 115 млн лет. Предположительный размер ископаемой рептилии был около 35 см, а размах её крыльев достигал 75 см (размер взрослой особи мог быть больше). Вероятно, Vectidraco — самый мелкий представитель всех Azhdarchoidea, так как другие известные самые мелкие аздархоиды (китайские Sinopterus и Huaxiapterus из семейства Tapejaridae) имеют размах крыльев 114 и 143 см соответственно. По мнению авторов описания, Vectidraco по своей общей морфологии и пропорциям тела был сходен с родом Tapejara и другими мелкими птерозаврами надсемейства Azhdarchoidea, был беззубым, возможно, с гребнем на голове, имел сравнительно короткие крылья и был способен к четвероногой локомоции. Вид был назван V. daisymorrisae в честь 9-летней девочки Дэйзи Моррис (Daisy Morris), обнаружившей останки рептилии на побережье острова. Она любила ходить по пляжу и собирать кости животных, ракушки, черепа и зубы. По мнению соавтора описания палеонтолога Мартина Симпсона из Университета Саутгемптона, куда в 2009 году обратились родители девочки, если бы юная исследовательница не нашла окаменелости, то они могли быть уничтожены и смыты морским прибоем. Название рода Vectidraco происходит от греческого имени острова Уайт (Vectis) и слова дракон («дракон с острова Уайт»). A scientific paper was published in 2013 about the find in the electronic journal PLoS ONE, titled A New Small-Bodied Azhdarchoid Pterosaur from the Lower Cretaceous of England and Its Implications for Pterosaur Anatomy, Diversity and Phylogeny. In it Darren Naish, Martin Simpson, and Gareth Dyke described and named the type species Vectidraco daisymorrisae. The generic name is derived from the Latin Vectis, the Roman name of the island now known as the Isle of Wight, and dracō, meaning "dragon". The specific name honours the discoverer Daisy Morris. A children's book has also been written by Simpson about Daisy Morris's discovery, called Daisy and the Isle of Wight Dragon. The only known specimen, holotype NHMUK PV R36621, was uncovered in the Chale Clay Member of the Atherfield Clay Formation of the Lower Greensand Group, a clay layer of the Deshayesites forbesi zone, Deshayesites fittoni subzone, dating from the early Aptian, with an age of 124 million years. It consists of the left side of a pelvis, the right ischium, the rear dorsal vertebra and the first three sacral vertebrae, of a subadult or adult individual. Vectidraco is a relatively small pterosaur. The pelvis is four centimetres long as preserved. Vectidraco's wingspan was estimated at seventy-five centimetres, its total body length at thirty-five centimetres. In view of its affinities, the describing authors assumed it was a toothless form, featuring a crest on its snout. Several unique derived traits, autapomorphies, were established. The hip joint is bordered at its top rear corner by a triangular depression. This depression is overhung by a ridge running downwards to the rear. The front blade of the ilium features an undivided roughly oval depression at its front inner side, below a convex surface. Furthermore a unique combination of traits is present in that the elongated rear blade of the ilium is T-shaped, terminating in a wide expansion also projecting upwards, that is longer than the shaft of the rear blade itself. Damage to the ilium shows the presence of camellate bone, internal air chambers. Also all the preserved vertebrae are pneumatised. Vectidraco was assigned to the Azhdarchoidea, in a basal position. If correct, this would make it one of the smallest azhdarchoids known ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| November 27th, 2014 | |
| 10:11 pm [industrialterro] [Link] |
Zhenyuanopterus Zhenyuanopterus is a genus of pterosaur which is known from Lower Cretaceous (early Aptian) Yixian Formation of Liaoning, China. It has been suggested that Zhenyuanopterus foraged while swimming, trapping prey within its needle-like teeth, a method similar to that of modern Platanista river dolphins, which display a similar dentition. This is a reasonably large pterosaur, with a wingspan of about 4 m (Lü 2010). One more interesting thing to say about these pterosaurs – and this isn’t unique to boreopterids but goes for ornithocheirids as well – look how tiny their feet are! It also seems that some ornithocheiroids lack fibulae, but it remains uncertain how widespread this is (among boreopterids, Boreopterus lacks them but this was assumed to be preservational, while Zhenyuanopterus has them). The enormous wings and very small, weak legs of many ornithocheiroids show pretty convincingly that they were predominantly aerial, that they walked little, and that they made a living by grabbing objects while in flight. What a strong contrast to long-legged, relatively short-winged pterodactyloids like the azhdarchids. It has been often suggested that this animal is actually the adult of its close relative Boreopterus, which is known to be a juvenile.
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, бореоптериды, диапсиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| 08:06 pm [industrialterro] [Link] |
Eurazhdarcho Eurazhdarcho is a genus of pterodactyloid pterosaur from the Late Cretaceous Transylvanian Basin of Romania. In 2009, Mátyás Vremir at Lancrăm near Sebeş-Glod in Transylvania at the SbG-B site uncovered the remains of a pterosaur. He donated these to the Erdélyi Múzeum, of the Societății Muzeului Ardelean (Transylvanian Museum Society). Subsequent excavations by Vremir discovered additional bones of the same individual animal and were added by him to the collection of the Babeș-Bolyai University. In 2013, Vremir, Alexander Wilhelm Armin Kellner, Darren Naish, and Gareth Dyke named and described the type species Eurazhdarcho langendorfensis. The generic name combines the name of Europe with that of the related form Azhdarcho. The specific name refers to Langendorf, the name of Lancrǎm in the language of the German ethnic minority in Romania. The article appeared in the electronic journal PLoS ONE without an accompanying printed version; it nevertheless validly names the taxon under the new rules of the ICZN. The holotype, EME VP 312, was found in a layer of the Sebeş Formation dating from the upper Early Maastrichtian, about 69 million years old. It consists of a partial skeleton lacking the skull. It includes three neck vertebrae among which the almost complete third and the fourth; the third and fourth right metacarpal; the upper part of the first phalanx of the wing finger; the lower part of the second phalanx; a lower phalanx of one of the other fingers and a number of undetermined fragments. The Babeș-Bolyai University material is included within this enumeration and is not indicated by a separate inventory number. Generally the quality of the bones is poor with much of the outer cortex broken or eroded and internal structures present as (impressions of) natural moulds. The fossils have not been completely flattened, preserving three-dimensionality, but compression has caused some distortion. The carcass had probably by flooding been deposed on its back in mud near a riverbank. Afterwards it was exposed to the air, weathering and being scavenged as proven by circular bite-marks inflicted by the conical teeth of some member of the Crocodyliformes. Later covered by a thin layer of dirt, it was damaged by beetles and termites. The authors noted that from the same Romanian layers the related giant form Hatzegopteryx is known; the known fossil material from both genera does not overlap. The authors considered an identity to be unlikely because the much smaller EME VP 312 seems to represent an adult individual. Eurazhdarcho is a medium-sized azhdarchid. The authors estimated its wingspan at three metres, extrapolating from an estimated length for the fourth metacarpal of about twenty-five centimetres. The authors established some distinctive traits, all present in the cervical vertebrae. The third neck vertebra has three-quarters of the length of the fourth vertebra, whereas 60% would be normal with azhdarchids. The necks of the prezygapophyses, the front joint processes, are well-developed and elongated, obliquely pointing forwards and outwards under an angle of 30° with the long axis of the vertebra. The preexapophysis, a secondary joint process on the side of the prezygapophysis, is well-developed with a forward pointing articulation facet and separated from a process, itself the remains of the diapophysis and possibly a neck rib, on the outer base of the prezygapophysis, by a deep trough on its underside. The pneumatic openings, the entrance holes for the air sacks, at the sides of the neural arch are small and placed in a low position. Eurazhdarcho was by the authors assigned to the Azhdarchidae, based on the method of comparative anatomy; a cladistic analysis was not performed. The area where Eurazhdarcho was found, in the Upper Cretaceous was localised on the Hațeg Island, part of the European Archipelago. The SbG-B site, though encompassing a surface of just 200 m³, has yielded several distinct animal species among which the turtle Kallokibotion bajazidi, the hadrosaur Telmatosaurus and a form referred to the titanosaur Magyarosaurus. This terrestrial fauna suggests that Eurazdarcho was not a coastal piscivore catching fish on the wing, affirming the "superstork" model for azhdarchids, in which they are terrestrial stalkers snatching small prey animals while walking on all fours. If Eurazhdarcho was indeed distinct from Hatzegopteryx, its discovery implies the presence of two azhdarchid forms in the Hațeg fauna, the one gigantic, the other medium-sized. This suggests a niche partitioning between them, although it is as yet unclear how this correlates with differences in prey preference and hunting techniques. This reflects a pattern seen in other Late Crataceous faunae which also show a combination of a large azhdarchid species with a smaller one. The Javalina Formation from the Maastrichtian of Texas has brought forth the giant Quetzalcoatlus northropi but also a smaller Quetzalcoatlus sp. and the azhdarchoid represented by specimen TMM 42489-2. The Two Medicine Formation fauna from the Campanian of Montana includes the smaller Montanazhdarcho minor, with a wingspan of 2.5 metres, but also fragments of larger forms with a span of eight metres. In the coeval Dinosaur Park Formation of Canada the smaller specimen RTMP 92.83 was discovered with a wingspan of five to six metres but also the large specimen PMA P.80.16.1367 indicating a ten metre wide animal. ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархиды, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| November 24th, 2014 | |
| 09:42 pm [industrialterro] [Link] |
Haopterus Haopterus is a pterodactyloid pterosaur genus from the Barremian-Aptian-age Lower Cretaceous Yixian Formation of Liaoning, China. It was in 2001 named by Wang Xiaolin and Lü Junchang. The type species is Haopterus gracilis. The genus name honours Professor Hao Yichun and combines his name with a Latinised Greek pteron, "wing". The specific name, "slender-built" in Latin, refers to the condition of the metatarsals. The genus is based on holotype IVPP V11726, a crushed fossil found in 1998 at the Sihetun-locality. The layer it was discovered in, was argon-dated at an age of 124.6 million years. It was the first Chinese pterosaur fossil preserving the skull. It consists of the front half of a subadult, including a skull, lower jaws, pectoral girdle, sternum, wings, cervical and dorsal vertebrae, partial pelvis and metatarsals. The skull is with a length of 145 millimetres long and low, lacking a crest. The snout is pointed but rounded. The maxilla and praemaxilla are completely fused with no visible suture. The nasopraeorbital skull opening is elongated and elliptical with a length of four centimetres, 27,6% of the total skull length. The lower jaws have a length of 128 millimetres. On the front two thirds of their length teeth are present. There are twelve pairs of teeth in both the upper and the lower jaws. The teeth are robust, sharp, pointed, and curving backwards. To the front they gradually increase in length and point more to the front. The first three pairs in the praemaxilla are very small though; the describers assumed these were replacement teeth, recently erupted. The back of the skull and the cervical vertebrae are strongly crushed, obscuring most details. Eight dorsal vertebrae are preserved, with a total length of 52 millimetres. The sternum was fan-shaped with a prominent keel. The wings are robustly built; the ulna is with 101 millimetres longer than the wing metacarpal which has a length of 89 millimetres. The wingspan of the type individual was estimated at 1.35 metre (4.43 ft). In comparison the hindlimbs must have been weakly built, the metatarsal having a length of just seventeen millimetres. The authors concluded that its slender hindfeet meant that it was forced to move quadrupedally on land, suggesting a piscivore lifestyle as a specialised soarer. Haopterus was by Wang classified as a member of the Pterodactylidae, mainly because of the combination of robust teeth with the lack of a skull crest. In 2006 however, a cladistic analysis by Lü showed it was a basal member of the Ornithocheiridae. Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейриды, орнитохейройды, птеродактили, птерозавры |
| November 23rd, 2014 | |
| 09:52 pm [industrialterro] [Link] |
Banguela Banguela oberlii is a dsungaripterid pterosaur from the Lower Cretaceous of Brazil. The Swiss collector Urs Oberli acquired a pterosaur jaw fragment from the Chapada do Araripe. In 2005, this was described by André Jacques Veldmeijer e.a. and referred to Thalassodromeus sethi. In 2006, Veldmeijer named it as a new Thalassodromeus species: Thalassodromeus oberli. However, this was done in his dissertation and thus merely resulted in an invalid nomen ex dissertatione. In 2014 it was named and described by Jaime Headden and Hebert Bruno Nascimento Campos as a separate genus Banguela, with the type species Banguela oberlii. The genus name is a Brazilian Portuguese word for "toothless one", especially used as an affectionate term for elderly women. The specific name honours Oberli. The holotype, NMSG SAO 251093, was probably found in the Romualdo Formation, also known as the Romualdo Member of the Santana Formation, dating from the Aptian-Albian. It consists of the symphysis, fused front end, of the lower jaws. Banguela has an estimated skull length of about two feet and wingspan of over twelve feet. The symphysis, with a preserved length of 273 millimetres, curves upwards and has a relatively short depression at its upper rear end. The front upper edge of the symphysis is sharp. The front bottom edge is sharp too but lacks a true crest. There are no teeth or tooth sockets present in the fragment. Veldmeijer had already in 2005 noted similarities to Dsungaripterus, but considered the available data to be insufficient to draw any conclusions from this. In 2014, Headden & Campos placed Banguela in the Dsungaripteridae, in a basal position. Banguela is unique among dsungaripterid pterosaurs due to a presumed total absence of teeth. Other pterosaur groups, such as pteranodontids, nyctosaurids and azhdarchoids, have also lost their teeth, indicating that toothloss might have independently occurred at least four times among pterosaurs. However, because dsungaripterids are occasionally recovered as derived azhdarchoids, it is possible that toothloss has occurred more often, if as an instance of Dollo's Law azhdarchoids should be originally toothless. If there was a large number of cases, Banguela suggests how it developed in most of these: the development of horned rhamphothecae in the jawtips, with progressive tooth rarification until they cease to be useful. It is worth to note that dsungaripteroids have some of the most specialised teeth of all sauropsids, so Banguela's toothlessness must indicate some degree of divergent specialisation.
Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, джунгариптероиды, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры |
| 08:09 pm [industrialterro] [Link] |
Ikrandraco Ikrandraco ("Ikran [a flying creature from Avatar with a crest on the lower jaw] dragon") is a genus of pteranodontoid pterosaur known from Lower Cretaceous rocks in northeastern China. It is notable for its unusual skull, which features a crest on the lower jaw. Ikrandraco is based on IVPP V18199, a partial skeleton including the skull and jaws, several neck vertebrae, a partial sternal plate, parts of both wings, and part of a foot. A second specimen, IVPP 18406, has also been assigned to Ikrandraco; it consists of a skull and jaws and the first three neck vertebrae. Both specimens come from the Aptian-age Lower Cretaceous Jiufotang Formation of Liaoning, with an estimated date of 120 million years ago. The type and only described species is I. avatar, a second reference to the movie Avatar. It was described in 2014 by Xiaolin Wang and colleagues. Ikrandraco is notable for having a very long, low skull (the height of the back of the skull, at the quadrates, is less than 19% the length of the skull), with a prominent blade-like crest on the underside of the lower jaw and no corresponding crest on the tip of the upper jaw, a crest combination not seen in other pterosaurs to date. The posterior edge of the crest also has a hook-like process. Each side of the upper jaw has at least 21 small cylindrical teeth, and each side of the lower jaw has at least 19. The skull of the type specimen is 286.5 millimetres (11.28 in) long, and the skull of the second specimen is at least 268.3 millimetres (10.56 in) long. Wang et al. performed a phylogenetic analysis including Ikrandraco and found it to be a basal pteranodontoid, more derived than Pteranodon but not as derived as the istiodactylids, anhanguerids, and other pteranodontoids. Wang et al. interpreted the crest as a possible adaptation for skim fishing, although they did not regard this as the animal's main method of foraging. The hook on the crest may have been an attachment point for a throat pouch for storing food, akin to a pelican. Ikrandraco was an approximate contemporary of distantly related anhanguerids Liaoningopterus gui and Guidraco venator, and all three are regarded as piscivores, but Ikrandraco differed from them in its much smaller and less robust teeth, indicating it had a different niche. ( Read More )</span>
Tags: Вымершие рептилии, Мел, авеметатарзалии, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеранодонтойды, птеродактили, птерозавры |
| 03:34 pm [industrialterro] [Link] |
Europejara Europejara is a genus of tapejarid pterosaur from the Early Cretaceous of Spain. In 2012, the type species Europejara olcadesorum was named and described by Romain Vullo, Jesús Marugán-Lobón, Alexander Kellner, Angela Buscalioni, Bernard Gomez, Montserrat de la Fuente and José Moratalla. The generic name combines the names of Europe and the related genus Tapejara, in reference to the fact that Europejara is the first tapejarid found in that continent. The specific name refers to the Olcades, the Celtiberic tribe inhabiting the region of Cuenca, the location of the find, in Antiquity. The holotype, MCCM-LH 9413, was uncovered at the Las Hoyas site in a chalkstone layer of the Calizas de La Huergina Formation dating from the late Barremian. It consists of a partial skull with lower jaws, compressed on a slab and counterslab. Two elements of the hyoid are present also. The skull has been vertically crushed, the lower jaws horizontally. The specimen was prepared by Mercedes Llandres Serrano, and is part of the Las Hoyas collection of the Museo de las Ciencias de Castilla–La Mancha. Europejara is a relatively small form with an estimated wingspan of two metres. The jaws are toothless and the lower jaws bear a large downwards pointing crest. The describers established three autapomorphies, unique derived traits. The crest on the lower jaws is curved to the back. The crest is deeper than its base, measured from the front to the back, is wide. The crest is four times deeper than the back of the jaw. Two other diagnostic traits were indicated: the inner side of the lower jaw is thickened, causing a convex curvature; the inner side shows some shallow, but well-demarcated, depressions. Due to the crushing of the skull, its fragments, mainly representing elements from the area around the right eye socket, show little detail. The lower jaws have a preserved length of twenty-three centimetres and an estimated original length of 255 millimetres. In their front parts the lower jaws are fused by a symphysis into a mandibula. The symphysis has a concave upper profile and features a large crest on the underside, pointing downwards for at least nine centimetres. The back edge of the crest is recurved; the curvature of the front edge cannot be exactly established because of damage. The crest is the longest relative to lower jaw length of any known pterosaur. The internal bone structure of the crest is spongy. The rod-like first ceratobranchialia pair of the hyoid have a length of 135 millimetres and a cross-section of two millimetres. Europejara was assigned to the Tapejaridae. A cladistic analysis showed it to be more precisely a member of the Tapejarinae. Apart from being the first tapejarid known from Europe, it would also be the oldest pterosaur with certainty known to be edentulous; older fragments have been reported representing other generally toothless clades but these did not include the jaws themselves. Following earlier suggestions about the diet of tapejarids, the describers assumed a frugivorous lifestyle for Europejara. Because the species is so old it indicates a rôle for the tapejarids in the Cretaceous Terrestrial Revolution, a turn-over in the ecosystems of the Lower Cretaceous in which gymnosperms were replaced by angiosperms, flowering plants, and new groups of herbivores evolved, adapted to the changed food supply. In the case of tapejarids there could have been a reinforcing interactive cycle between the evolution of fruit and the pterosaurs dispersing the seed. Possibly the beaks of the tapejarids had ragged edges forming pseudo-teeth to better separate the fruit flesh from the seeds, as with some extant toucans. ( Read More ) Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры, тапежариды |
| 01:43 pm [industrialterro] [Link] |
Caiuajara Caiuajara is an extinct genus of tapejarid pterosaur from the Late Cretaceous of Brazil. It is known from a single type species, Caiuajara dobruskii. In 1971, the labourers Alexandre Dobruski and his son João Gustavo Dobruski found pterosaur fossils in a field near Cruzeiro do Oeste in the south of Brazil, in the state of Paraná. The finds were in 2011 brought to the attention of paleontologists Paulo C. Manzig and Luiz C. Weinschütz. In 2014, the type species Caiuajara dobruskii was named and described by Paulo Manzig, Alexander Kellner, Luiz Weinschütz, Carlos Fragoso, Cristina Vega, Gilson Guimarães, Luiz Godoy, Antonio Liccardo, João Ricetti and Camila de Moura. The generic name refers to the geological Caiuá Group and the related genus Tapejara. The specific name honours the discoverers. The holotype, CP.V 1449, was found in a sandstone layer of the Goio-Erê Formation, the age of which is uncertain; it is perhaps dating from the Coniacian-Campanian, very roughly about eighty-five million years old. It consists of a partial skeleton including the skull, lower jaws, neck vertebrae and wing elements. Many hundreds of bones have been discovered, concentrated in several bone beds, and representing at least forty-seven individuals but probably many more. In the total assembly, all elements of the skeleton are present. The bones have been three-dimensionally preserved, not compressed, but are only rarely articulated. The individuals found are often juveniles; adult animals are much rarer, only represented by two skulls and three humeri. Good specimens have been assigned as paratypes, the more fragmentary ones have been referred. The paratypes are: CP.V 865: a snout, rear of the mandibula, right jugal, vertebrae, ribs and metatarsals; CP.V 867: a snout and limb bones; CP.V 868: a snout, wing elements and other postcrania; CP.V 869: a vertebral column, right arm, coracoid, breastbone, wing phalanges, belly ribs, pelvic elements and a right tighbone; CP.V 870: a shoulder girdle with the humeri; CP.V 871: a right shoulder girdle with right arm elements; CP.V 872: s partial skeleton including the skull, lower jaws, right arm, neck vertebrae and additional limb elements; CP.V 873: a snout and finger phalanges; CP.V 999: a partial skull; CP.V 1001: a slab with a partial skull, lower jaws and postcrania of at least three individuals; CP.V 1003: a partial skull and symphysis; CP.V 1004: a snout; CP.V 1005: a partial crested skull with the complete mandibula; CP.V 1006: a partial crested skull lacking the snout combined with postcrania; CP.V 1023: a snout and postcrania; CP.V 1024: a skull and postcrania of at least three juveniles; CP.V 1025: a thighbone; CP.V 1026: a thighbone; CP.V 1450: a slab containing at least fourteen juveniles; CP.V 2003: a skull with lower jaws and articulated wing elements; UEPG/DEGEO/MP-4151: a slab with two skulls and postcrania; and UEPG/DEGEO/MP-4152: a snout with postcrania. Most specimens are part of the collection of the Centro Paleontológico of the Universidade do Contestado. The largest individuals of Caiuajara had an estimated wingspan of 2.35 metres. The species had a large toothless head with, in adult individuals, an enormous shark fin-shaped crest on the snout. The describing authors established several distinguishing unique traits, autapomorphies. The tip of the snout is strongly oriented to below, at 142 to 149°, relative to the edge of the upper jaw. The rear ascending branches of the praemaxillae on their midline form an elongated bony rim projecting to below into the nasoantorbital fenestra, the large skull opening in the side of the snout. In the concave upper rear of the symphysis, the fronts of the lower jaws grown together, a rounded depression is present. The front outer edge of the quadrate shows a longitudinal groove. Below the front part of the nasoantorbital fenestra, a depression is present in the upper jaw edge. Additionally, Caiuajara shows an unique combination of traits that are themselves not unique. The lower edge of the eye socket is rounded. At a maximal occlusion, the gap between the upper and lower jaw is wider than with other tapejarines. The pteroid on its bottom surface shows a conspicuous depression lacking a pneumatic opening. Caiuajara was assigned to the Tapejaridae, more precisely the Tapejarinae. It shares several traits with the tapejarids, such as a crest running from the front snout to the back of the head; an elongated nasoantorbital fenestra occupying over 40% of total skull length; and a large boss on the front edge of the coracoid. A typical tapejarine trait is the down-turned snout tip. A cladistic analysis showed that Caiuajara is a possible sister species of Tupandactylus. In 2014, Caiuajara was the geologically youngest known tapejarid (aside from the possible tapejarid Bakonydraco galaczi) and also the most southern one known. This expansion of their known range was seen as an indication that tapejarids had a global distribution. Moreover, Caiuajara is the first pterosaur found in the south of Brazil. The habitat of Caiuajara was a desert with dunes. The layers in which the fossils were found had been deposited in a lake in the desert; probably the bones had been exposed at the surface around the lake for a time and were then by storms blown into it, eventually sinking to the bottom. Possibly the same storms caused many individuals to die together; this could also have been the result of droughts. A succession of layers shows that the lake was likely inhabited by the pterosaurs for a great length of time, although it is also possible they visited the lake during regular migrations. Fossil plants — tapejarids are often assumed to have been herbivores — have not been found, so there are no direct indications about the food source. Likewise, remains of invertebrates have not been discovered. The large concentrations of fossils, among pterosaurs very rare and only equalled by those found of the Argentine form Pterodaustro, were by the describers seen as proof of a gregarious lifestyle, Caiuajara living in colonies. The many specimens also allowed to determine a growth series, the first such an ontogenetic sequence for pterosaurs of which it is nearly certain that it really represented a single species. The age of the exemplars can be determined, not just from size but also by the degree of ossification, especially of the breastbone, the long bones and the wrist, and the fusion of the shoulder blade and coracoid into a scapulocoracoid. It showed that juvenile individuals, the smallest specimens of which have a wingspan of about sixty-five centimetres, generally had the same proportions as adults. Especially important is that their humeri are not proportionally smaller and their humeral deltopectoral crests, the attachments of the main flight muscles, are not less developed, attaining a size of 38 to 40% of the humeral shaft length. This suggests that they were precocial, taking wing almost as soon as they hatched; parental care must have been limited. This might have been typical of all derived pterosaurs. The snout crest however, strongly changed during growth. It became much taller and also much more steeply inclined, from about 115° to 90°. Although the snout as a whole also became more massive, the snout tip inclination relative to the jaw edge remained the same. At the back of the skull an additional projection developed. Furthermore the dentary crest on the lower jaw strongly increased in size. No specimens have been found lacking the snout crest, indicating that Caiuajara was in this respect not sexually dimorphic and casting doubt on the hypothesis that pterosaurs normally were. ( Read More ) Ископаемые останки (1, 2, 3, 4, 5):
Tags: Вымершие рептилии, Мел, авеметатарзалии, аждархойды, архозавроморфы, архозавры, диапсиды, монофенестраты, орнитохейройды, птеродактили, птерозавры, тапежариды |



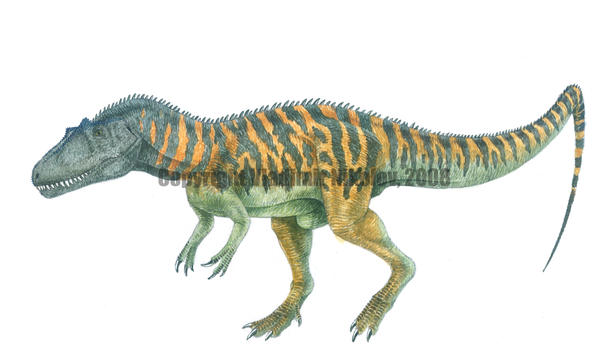


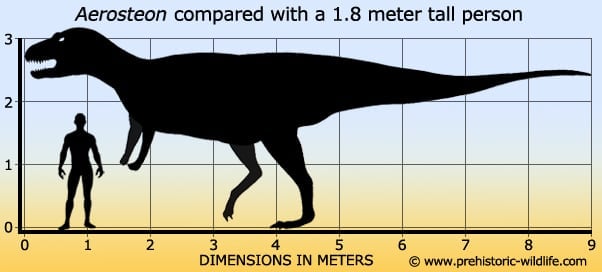




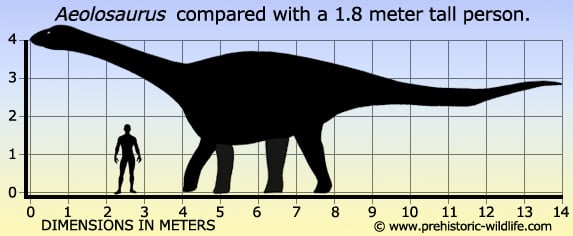



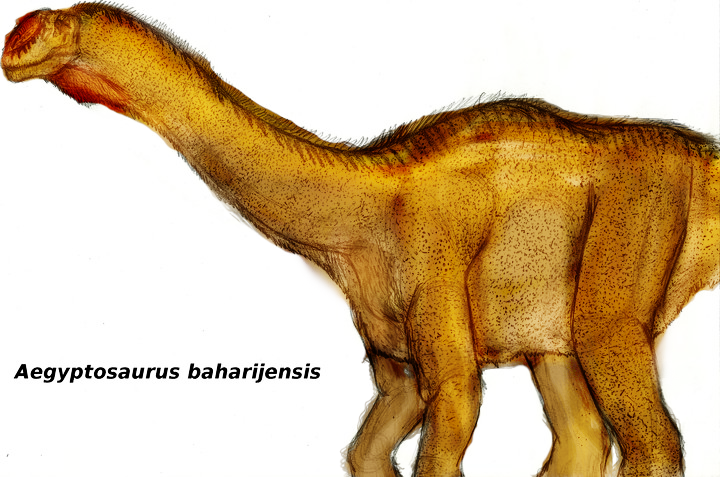
















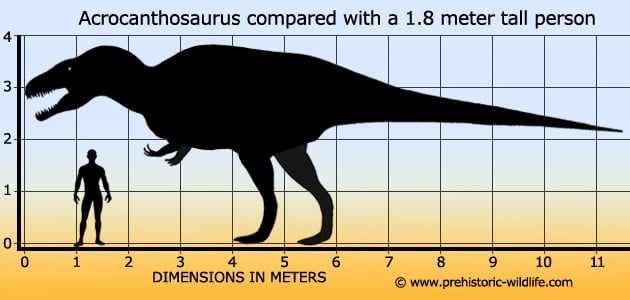



















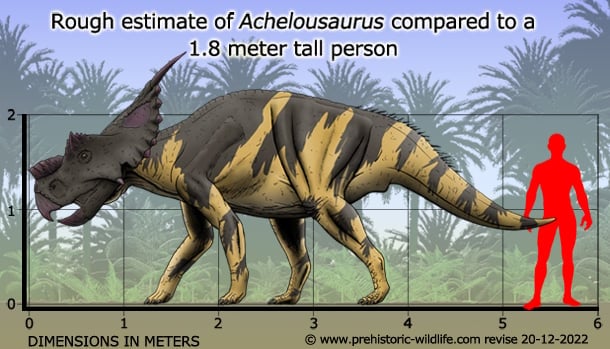




















































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg?uselang=ru){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}